La sémiotique des mondes vivants
Du signe à l’interaction, de la téléologie à la structure
Jacques Fontanille
Centre de Recherches Sémiotiques
Université de Limoges
Index
Mots-clés : biosémiotique, interactions, téléologie, Umvelt
Auteurs cités : ARISTOTE, Francis BACON, Marcelo BARBIERI, Denis BERTRAND, Bruno CANQUE, Charles DARWIN, Richard DAWKINS, Jacques DEWITTE, Jacques FONTANILLE, GROUPE µ, Hans JONAS, Jean-Marie KLINKENBERG, Kalevi Kull, Eric LANDOWSKI, Maurice MERLEAU-PONTY, Howard PATTEE, Adolf PORTMANN, Bernardin de SAINT-PIERRE, Robert SPAEMANN, Jacob von UEXKÜLL, Thure von UEXKÜLL, Larry WRIGHT
1. Introduction
L’expression « monde vivant », notamment au pluriel (mondes vivants), est ici un équivalent d’Umwelt (Umwelten). Nous réservons la notion d’« univers » à ce que Greimas désigne et définit dans Sémantique structurale comme des « univers de sens », c’est-à-dire comme les domaines où s’exerce la perception sémantique des discontinuités sur lesquelles se fonde la construction de la signification. Les mondes vivants sont des domaines d’interaction, centrés sur une entité vivante dont l’observateur adopte le point de vue. Un micro-univers de sens peut certes coïncider avec un monde vivant, mais alors que dans le premier cas c’est l’observateur en tant que tel qui se place au centre des effets de sens perçus, dans le second, c’est un organisme vivant quelconque qu’on trouve au centre des interactions.
- Note de bas de page 1 :
-
Groupe μ, Principia semiotica. Aux sources du sens, Bruxelles, Les Impressions Nouvelles, 2015.
Dans Principia semiotica, le Groupe μ vise une sémiotique générale qui intègrerait la signification du monde vivant en général1. Jean-Marie Klinkenberg, l’un des deux auteurs, s’interroge par exemple sur ce que serait le régime de signification propre à un ver de terre :
- Note de bas de page 2 :
-
Jean-Marie Klinkenberg, cité en ligne dans « Pourquoi y a-t-il du sens plutôt que rien ? », Julia Luong, 2018, Université de Liège - http://www.reflexions.uliege.be/cms/c_420022/en/pourquoi-y-a-t-il-du-sens-plutot-que-rien?printView=true - September 23, 2018.
Le ver de terre meurt s’il est exposé à la lumière. Or si vous l’y exposez, il va tenter de rentrer dans le sol : est-ce que cela veut dire qu’il est conscient ? Non. Mais le ver de terre est néanmoins en possession du minimum de ce qu’est un langage, c’est-à-dire un plan d’expression et de contenu. Sur le plan de l’expression, on trouve ici l’opposition clair/sombre, qui existe parce qu’il y a chez le ver de terre des capteurs qui lui permettent de la manipuler, de la gérer et de l’exagérer. Sur le plan du contenu, on trouve l’opposition correspondante vie/mort. Le fonctionnement du ver de terre repose donc sur des informations organisées selon un système d’oppositions binaires. Et c’est un système de sens qui lui permet de survivre. Bien sûr, c’est une grammaire extrêmement simple, que le vers de terre n’est pas capable de décrire, contrairement à nous. Néanmoins, à bien y réfléchir, peu de gens sont capables de dire comment fonctionne leur langue.2
Cette extension des investigations sémiotiques n’est pas nouvelle, certes, puisque la sémiotique d’inspiration peircienne, notamment aux Etats-Unis et en Europe du Nord, développe depuis le milieu du XXe siècle le domaine de recherche biosémiotique. En revanche, les sémiotiques structurales ont longtemps tenu le monde vivant hors de leur champ, principalement en raison d’une orientation naturaliste (au sens où l’entendent Philippe Descola et les anthropologues contemporains) qui a conduit les recherches d’inspiration structurale à se développer dans la direction d’une sémiotique des cultures, étant entendu que la signification de la nature pouvait être également appréhendée, mais seulement à travers le filtre des cultures.
Or il est évident que si le couplage semi-symbolique entre l’opposition vie / mort et l’opposition clair / sombre a un « sens » pour le ver de terre, ce n’est pas à travers le filtre d’une culture, sauf à supposer — ce qui n’est pas exclu — que n’importe quel être vivant construit un monde signifiant grâce à ses interactions avec son proche environnement. Mais le mouvement d’expansion qui porte l’ouvrage Principia semiotica ne s’en tient pas au vivant, car le Groupe μ tient le même type de raisonnement pour les machines, et même pour des dispositifs techniques relativement simples comme une soupape de sécurité ou un régulateur de Watt. Une machine en effet, aussi simple soit-elle, interagit elle aussi à sa manière avec son environnement en y sélectionnant des propriétés pertinentes et en ignorant toutes les autres.
Nous nous interrogeons ici sur le type de sémiotique dont nous avons besoin pour construire une biosémiotique qui serait d’inspiration structurale. Par conséquent, même si nous devons avoir le souci de ne jamais rompre les liens avec une sémiotique du monde humain, d’une part, et une sémiotique générale qui dépasserait le monde vivant, d’autre part, il n’en reste pas moins que nous devons comprendre ce qui est propre à la vie dans le monde vivant. Sinon, il n’est nul besoin d’une biosémiotique, et une sémiotique générale suffirait, comme le propose le Groupe μ.
- Note de bas de page 3 :
-
Jacob von Uexküll, Milieu animal et milieu humain, Paris, Payot, Bibliothèque Rivages, traduction de Charles Martin-Fréville, introduction de Dominique Lestel, « De Jacob von Uexküll à la bio-sémiotique », 2015 [2010]. — Sur l’éthosémiotique de von Uexküll, on pourra consulter une présentation plus détaillée et extensive in J. Fontanille, « La sémiotique des interactions chez von Uexküll », in Alessandro Zinna (éd.), La Sémiotique et les Sciences. Biologie, Éthologie et Sémiotique. Les vivants et leur environnement. Milieu, habitat, territoire, espace familier, Actes du colloque d’Albi-Moissac 2018, à par.
- Note de bas de page 4 :
-
Nicole Pignier, dans « Design et éco-sémiotique. Quand le design co-énonce avec le vivant » prend à cet égard les précautions nécessaires : en faisant appel à l’énonciation, elle précise que l’actant qui en serait responsable n’est ni un sujet ni un actant transcendantal, mais un « actant ambiant », une « tension actantielle entre instances partenaires accueillants / accueillis » (p. 72). (Dans E. Mitropoulou et N. Pignier, Le sens au cœur des dispositifs et des environnements, Paris, Connaissances nouvelles, 2018).
Jacob von Uexküll s’est lui-même posé la question en faisant référence à Descartes : dans son introduction à l’ouvrage Milieu animal et milieu humain, il précise qu’à la différence de Descartes (pour qui les animaux sont tout comme des machines), il va remettre le machiniste dans la machine3. Ce geste inaugural ne consiste pas seulement à identifier un actant dans la machine, puisque la machine en est déjà un, mais à introduire un actant à l’intérieur de l’actant : le « machiniste » est un actant interne, un pilote ou un centre de contrôle qu’Uexküll appelle sans doute trop rapidement un « sujet »4. Néanmoins, c’est bien cette opération qui l’autorise à concevoir un Umvelt, car le domaine d’interactions d’un organisme vivant doit être organisé autour d’un centre de référence, qui fait toute la différence avec l’environnement quelconque d’une machine et de quelque dispositif technique que ce soit.
- Note de bas de page 5 :
-
Nous conservons ici systématiquement le terme allemand, car aucune traduction française n’est satisfaisante. « Milieu », qui est souvent utilisé en traduction, est pourtant un concept zoologique qu’Uexküll récuse : dans la tradition lamarckienne, et même pour le sens commun, le milieu est un ensemble de déterminations qui s’imposent aux êtres vivants, alors que l’Umwelt est un domaine d’interactions co-construit par l’organisme et le milieu.
- Note de bas de page 6 :
-
Dans un but et avec des arguments comparables, Augustin Berque a choisi le terme de « subjectité » (Augustin Berque, Ecoumène. Introduction à l'étude des milieux humains, Paris, Belin, 2009 (2000). Ce terme et cette notion sont empruntés à Emmanuel Levinas, Autrement qu’être ou au-delà de l’essence, Paris, Livre de Poche, 1990 [1974], p. 134.
Le machiniste en question est bien loin d’être un « sujet », quelle que soit l’acception que l’on donne à ce terme. C’est une instance actantielle, certes, qui perçoit, ressent, contrôle et agit, mais qui n’est ni sujet, ni objet : partenaire d’un ensemble d’interactions, elle n’est qu’un interactant parmi d’autres. Ce sera donc notre hypothèse de travail : les mondes vivants sont des domaines d’interactions entre chaque organisme et chaque espèce et leur milieu, organisés autour d’un centre actantiel de référence, qui est l’un des interactants dont on adopte le point de vue pour construire son Umwelt5. Pour éviter d’inévitables confusions, nous parlerons à cet égard de subjectalité et de non de subjectivité : la subjectalité en question n’est en l’occurrence que le nom de l’orientation épistémique que, par principe de méthode, on adopte pour spécifier et différencier les mondes vivants de tous les autres6.
- Note de bas de page 7 :
-
Hans Driesch, Geschichte des Vitalismus, Whitefish, Kessinger reprints, 2010 [1905].
- Note de bas de page 8 :
-
Chez Aristote, l’entéléchie est à la fois le processus et son résultat, dans ce qu’ils ont de plus parfaitement achevé, complet. En somme, l’actualisation ultime de la puissance.
La subjectalité est une hypothèse de travail (nous verrons même qu’elle peut être réduite à un effet de sens de l’actualisation des Umwelten), en ce sens qu’elle nous oriente vers un choix de méthode susceptible de nous donner accès aux principes de structuration du monde vivant. Ce choix de méthode ne présuppose en aucune manière une autonomie (ontologique) de la vie, et par conséquent ne peut être mis au crédit d’une conception qui se rapprocherait de l‘organiscisme ou du vitalisme. A la fin du XIXe siècle, Hans Driesch défendait par exemple l’existence d’une force vitale7 qui lui fournissait l’argument majeur pour fonder l’autonomie de la vie, étayée par ailleurs sur le concept aristotélicien d’entéléchie8. Toutes les manifestations éventuelles de cette « force vitale » étant indirectes ou incertaines, cette conception se vit opposer de nombreuses réfutations, la principale étant qu’elle substituait à des faits tangibles une explication inaccessible à quelque observation que ce soit. En revanche, la subjectalité n’étant que l’effet d’une orientation épistémique de l’Umwelt, sans laquelle la structuration et la signification des mondes vivants resterait inaccessible à la connaissance, elle ne se confond pas avec elles. En résumé : ne confondons pas ce qui nous permet de connaître quelque chose (des conditions épistémiques) avec les propriétés ontologiques de cette chose !
2. Le « but » de la vie : critique de la téléologie biosémiotique
- Note de bas de page 9 :
-
Comme nous avons par ailleurs développé en détail cette critique de la téléologie, nous renvoyons le lecteur à Jacques Fontanille, « Pour une biosémiotique des interactions. Régimes de signification et téléologie », in Louis Hébert et Stéphanie Walsh-Mathews (éds.), Points aveugles et points borgnes de la sémiotique, Semiotica, 2019.
Parmi toutes les dérives plus ou moins « métaphysiques » qui guettent ou tentent la biosémiotique, l’une d’entre elles domine, sinon par sa force explicative, du moins par sa récurrence dans les travaux des spécialistes du domaine : la téléologie, c’est-à-dire l’explication de la vie par les finalités du processus et des formes qu’elle adopte9. La téléologie pourrait être examinée a minima à l’aune de notre questionnement : de quelle sémiotique avons-nous besoin ? la téléologie est-elle une des explications requises pour traiter du vivant ? Eclaire-t-elle plus qu’elle n’obscurcit, et vice-versa ? Mais l’une de ses particularités, qui nous incite à un surcroît de précautions, est d’entraîner avec elle d’autres positions, qui sont, elles, et sans aucun doute, des erreurs de méthode : la projection d’explications ou de descriptions anthropomorphes sur les mondes animaux et végétaux, une hiérarchisation des espèces qui place l’espèce humaine en position dominante, le recours à une entité explicative invérifiable (un démiurge, un deus ex machina, un dessein intelligent, etc.).
2.1. La critique de la téléologie
2.1.1. Résistance à la vérification / falsification
La plupart des critiques adressées à l’explication téléologique, dans l’histoire de la pensée occidentale, portent justement ces positions associées à la téléologie : l’anthropomorphisme (critique portée notamment par Leibniz), le recours à un démiurge, un « actant caché » dans la machine (Aristote contre Platon), ou même l’obscurantisme (Spinoza dénonçant l’explication téléologique comme ignorance des « vraies » causes).
- Note de bas de page 10 :
-
Francis Bacon, Novum organum (1620), Paris, PUF, 1986.
Mais les critiques directement adressées à la téléologie elle-même sont plus récentes, et moins répandues : elles sont alors inspirées par les principes de la méthode scientifique. Francis Bacon, par exemple, dans le Novum Organum, s’attache à montrer que l’explication téléologique est une dérobade, car au mieux elle dispense de la vérification expérimentale, et au pire, elle la rend impossible10. L’argument est simple : une finalité (et surtout si elle est conçue sur le modèle de l’entéléchie aristotélicienne — complète, parfaite, accomplie) n’offre aucune possibilité de confrontation avec des faits, des manifestations observables qui sont toujours, par définition, provisoires, imparfaites et en métamorphose. Une finalité n’étant pas même une prévision ou une prédiction qui pourraient, lors d’une échéance déterminée ou estimée par anticipation, être confrontée à ce qui sera effectivement réalisé, elle n’est qu’une projection et une ligne de fuite eschatologique. Si elle pouvait être confrontée aux faits, elle ne serait pas plus qu’une condition (et non une finalité), éventuellement nécessaire à l’advenue de ces faits.
Cette critique concerne la sémiotique, au moins pour deux raisons. Tout d’abord, l’analyse sémiotique la plus élémentaire s’efforce de repérer des transformations : inflexions, métamorphoses, inversions de contenus, apparitions et disparitions. Et pour cela, elle doit s’appuyer, empiriquement, sur des manifestations observables (par exemple les deux formes successives d’une figure, ou le remplacement d’une figure ou d’une situation par une autre), qui lui permettront ensuite de construire une relation entre des expressions et des contenus. Or une explication téléologique est quasiment par définition un contenu sans expression : si le telos avait une expression, ce serait en effet seulement une prédiction ou une prévision (cf. supra). Pour l’analyse sémiotique, un projet ou un but sont nécessairement exprimables ou exprimés, hic et nunc, et peuvent de ce fait même appartenir à une sémiotique-objet précise (par exemple à un discours qui expose le projet ou le but).
Ensuite, quand la sémiotique s’intéresse au vivant, elle dialogue inévitablement avec la biologie, et par conséquent avec une pratique scientifique qui ne peut se soustraire aux principes de reproductibilité des protocoles de recherche, et de vérification / falsification. Ce dialogue est indispensable pour que les phénomènes qu’elle explore soient au moins scientifiquement valides, et régulièrement actualisés en fonction des avancées de la recherche, par la communauté des chercheurs qui les établit en tant que faits. Certes, la sémiotique est en droit de défendre l’autonomie de son objet, la signification, mais cette autonomie n’est qu’une position épistémologique et méthodologique, qui ne suffit pas à entraîner l’adhésion des biologistes aux analyses qui en découlent. Or l’explication téléologique est précisément de celles qui dispensent du recours aux faits et à leur validation, et qui remettent en cause la méthode scientifique en biologie. La biosémiotique est-elle en mesure d’assumer la rupture du dialogue qui s’ensuit ? Dispose-t-elle d’un modèle d’explication global et alternatif, reposant sur la postulation d’un actant caché et des intentions qu’il porterait ? Ce modèle éventuel serait-il susceptible de produire des connaissances sur le vivant qui seraient opposables à celles que produit la biologie, et de convaincre lors d’inévitables controverses scientifiques ? Rien n’est moins sûr.
- Note de bas de page 11 :
-
Charles Darwin, L’Origine des espèces, trad. Thierry Hoquet, Paris, Seuil, 2013.
C’est la raison pour laquelle, au nom de la méthode scientifique, la téléologie est le plus souvent renvoyée à la métaphysique, et exclue du champ de la connaissance scientifique. L’argument de Darwin est pourtant encore d’une autre nature11. Il s’efforce en effet de dissocier les changements opérés par la sélection naturelle de quelque « intention » directrice que ce soit, de toute force préalable qui serait inhérente au monde de la vie. Ce n’est visiblement pas en raison d’une hostilité à l’égard des intentionnalités directrices, mais plutôt d’un souci bien compréhensible d’aménager aussi largement que possible un espace théorique et méthodologique pour la sélection elle-même. Pour Darwin, l’évolution est faite d’événements prévisibles ou imprévisibles, probables ou aléatoires (donc modalisés) mais elle ne suppose aucune force cachée, aucun « actant » qui guiderait par exemple vers l’adaptation à un milieu ou vers la diffusion d’une espère.
Tout repose en effet sur un processus de tri appliqué à des séries d’« accidents ». Le tri lui-même n’est imputable à aucun actant interne ou externe, susceptible d’être manifesté par des acteurs identifiables, car la sélection n’est constatée qu’après-coup, à partir des innovations durables qu’elle produit : ces innovations existent, les solutions éliminées n’existent pas (ou plus), et la seule chose que nous puissions constater, c’est que la vie persiste, c’est que le cours de l’existence des mondes vivants n’est pas interrompu, et que cette persistance est à mettre au crédit des transformations que les mondes vivants ont traversées. On constate une transformation, on peut présupposer un opérateur de cette transformation, donc un actant, mais cet actant est une place vide, un pur opérateur sémiotique dont la manifestation est indéfiniment repoussée. A la limite, seule la disparition de la vie, l’interruption du cours d’existence, permettrait éventuellement d’identifier les manifestations de cet actant.
Darwin désactantialise l’évolution (pas de forces cachées ou de directions intentionnelles), mais sans la démodaliser (il manipule des pouvoir-être et des savoir-être, le prévisible et l’imprévisible, des probabilités, des impossibilités et des aléas). Cette nuance est, d’un point de vue sémiotique, décisive, car dans la sémiotique greimassienne et post-greimassienne, celle qui s’est le plus précisément consacrée à la structure actantielle de la signification, la théorie des modalités était destinée à remplacer en partie la théorie actantielle, et notamment dans toutes les situations d’analyse où il conviendrait d’éviter des concepts anthropomorphes comme « sujet », « objet », « destinateur », etc. Par conséquent, même sans actant identifiable auquel on puisse l’imputer, le processus de l’évolution peut être modalisé, et ces modalisations seront le support d’une possible construction de sa signification.
Enfin, la version la plus récente de ces critiques et des réinterprétations de la téléologie est l’œuvre de la cybernétique. La cybernétique distingue en effet la téléologie proprement dite, qui implique une intention, un site de l’intention, un actant caché, en somme une intentionnalité spéculative, et la téléonomie, qui implique seulement une nécessité associée en toutes ses phases à un processus, ou comme l’écrivait Kant à propos du télos, une « idée régulatrice ». Pour rendre compte des processus de commande, de communication et de régulation dans le monde du vivant (mais aussi dans les machines et dans les systèmes sociologiques et économiques), la cybernétique distingue les « systèmes gouvernants » (ou systèmes de contrôle) et les « systèmes gouvernés » (ou systèmes opérationnels). Elle procède par traduction du langage de description des systèmes de contrôle dans le langage cybernétique des systèmes opérationnels : au cours de cette traduction, les finalités proposées dans le premier sont converties en lois de régulation dans le second.
- Note de bas de page 12 :
-
Voir notamment Robert Nadeau, Sur la légitimité du langage téléologique en science biologique, conférence en ligne, UQAM, 1984 (https://unites.uqam.ca/philo/profs/nadeau/textes/Langage_teleologique1984.pdf).
En effet, dans le rapport entre systèmes gouvernants et systèmes gouvernés, la téléologie dirigerait et induirait, alors que la téléonomie oriente et régule seulement : ce serait en quelque sorte un degré faible de l’intentionnalité. La téléonomie oriente par exemple les organismes vivants vers toujours plus de complexité, étant entendu que cette complexité, jusqu’à un certain seuil, garantit leur identité, leur autonomie (leur « self-maintenance »), et donc leur persistance12. La solution cybernétique est séduisante : en répartissant les finalités et les lois de régulation sur deux dimensions différentes des systèmes analysés, elle rend possible l’affaiblissement du lien de détermination entre les deux, de sorte qu’on peut proposer une méthode de description des lois de régulation sans faire appel aux explications téléologiques, c’est-à-dire aux finalités.
La nécessité et la régulation téléonomique ont pourtant encore, sinon vraiment un but, du moins une orientation déterminée vers la complexité et l’autoconservation non seulement du vivant en général, mais aussi de chaque être individuel et surtout de chaque espèce. La régulation des interactions entre les deux types de systèmes, notamment en raison des rétroactions qui valident l’augmentation de la complexité, n’échappe donc pas complètement au raisonnement téléologique : ces complexifications autoproduites par les systèmes gouvernés ne sont prises en considération et validées par les systèmes gouvernants que dans la mesure où elles satisfont les finalités de ces derniers. En d’autres termes, la nécessaire isomorphie entre les deux types de systèmes, y compris obtenue par régulation, et transposée en deux langages inter-traductibles, légitime indirectement les visées téléologiques même reformulées. La cybernétique euphémise en quelque sorte les « comportements intentionnels des systèmes » en les qualifiant de téléonomiques.
Nous retrouvons ici les limites d’un raisonnement qui confond le fait même d’exister et le but de l’existence. En effet, puisque l’argument principal de la cybernétique en faveur de la téléonomie réside dans la « self maintenance » des organismes (qui changent en conservant leur identité, et donc en persistant), elle vise une condition, peut-être même « la » condition pour que les mondes vivants persistent à exister. Mais comme il n’y a pas d’existence sans persistance, cette explication est tautologique, et la condition en question n’est pas une explication de la persistance, mais bien la seule forme sémiotique que peut prendre l’existence, à savoir celle des organisations syntagmatiques du cours d’existence qui sont telles que nous pouvons dire « ça existe et ça continue d’exister ». La signification de l’existence ne se loge pas en effet dans ses conditions ontologiques, mais dans les formes syntagmatiques de la persistance.
- Note de bas de page 13 :
-
Jacques Monod, Le hasard et la nécessité, Paris, Seuil (coll. Points), 1973. Henri Atlan, Entre le cristal et la fumée, Paris, Seuil, 1979.
A la limite, pour Jacques Monod, ou Henri Atlan, les finalités deviennent des illusions suscitées par les effets du hasard, et notamment par la capacité des systèmes vivants à accumuler et conserver ces effets : cette capacité n’est pas en effet pour eux une « finalité », car elle est seulement ce qui est nécessaire pour que le vivant persiste13. En revanche, pouvoir mémoriser et accumuler les effets d’événements aléatoires est une propriété spécifique des mondes vivants, sur laquelle peut porter l’interprétation, et dont la sémiotique peut interroger la signification.
2.1.2. L’interprétation requise est risquée
Au moment d’exposer les résultats de la recherche, la pratique scientifique doit les interpréter pour les confronter aux connaissances existantes et dégager ainsi l’apport spécifique d’un protocole expérimental et/ou d’une argumentation. Par conséquent, la pratique scientifique opère au moins une transposition ou une traduction entre deux configurations sémiotiques : celle de l’expérience, de l’observation ou de l’argumentation, d’une part, et celle de leur interprétation, d’autre part. Les langages de l’interprétation peuvent être verbaux, visuels, ou autres, mais quels qu’ils soient, ils n’échappent pas aux contraintes, aux conséquences et aux aléas de la transposition intersémiotique. C’est notamment le cas (le plus fréquent) quand la sémiotique de l’interprétation est verbale. Ce serait donc la « couche sémiotique » de la pratique scientifique, dont aucune science ne peut se défaire, qui serait responsable des dérives téléologiques. Pour la biologie notamment, chaque concept verbalisé, chaque mot peut être un piège téléologique.
C’est précisément (et paradoxalement) ce piège verbal qui constitue l’argument principal des biosémioticiens qui défendent une perspective téléologique. Examinons par exemple la critique adressée naguère à la biologie « asémiotique » (et d’inspiration darwinienne) par un collectif de biosémioticiens :
- Note de bas de page 14 :
-
Notre traduction. Kalevi Kull, Terrence Deacon, Claus Emmeche, Jesper Hoffmeyer, Frederik Stjernfelt, « Theses on Biosemiotics : Prolegomena to a Theoretical Biology », Biological Theory, June 2009, vol. 4, 2, p. 169.
La biologie néo-darwinienne pratiquée dans le monde entier a adopté par défaut (c’est-à-dire, extrait du support contextuel nécessaire) une conception asémiotique de la vie comme une simple chimie moléculaire, et en même temps, elle dépend d’hypothèses sémiotiques non analysées. La raison pour laquelle ce n’est pas ressenti comme un problème est que la biologie compense l’exclusion de la sémiose en introduisant une pléthore de termes implicitement sémiotiques comme « information », « adaptation », « signal », « repère », « code », « messager », « fidélité », et « communication aléatoire ». Ces utilisations sont rarement bien définies et sont souvent appliquées de manière prétendument métaphorique, avec l’hypothèse implicite qu'ils peuvent être réduits à de simples opérations chimiques si nécessaire. Il n’est pas clair, cependant, qu’une réduction complète de ce genre est possible sans poser de problèmes. Si on demandait aux biologistes d’éviter ces termes implicitement sémiotiques, il leur serait difficile — et probablement impossible — d’expliquer la nature de la fonction organique.14
La critique adressée à la biologie repose entièrement sur la terminologie dont elle use : grosso modo, les biologistes darwinistes (il existerait donc des biologistes anti-darwiniens ?), obligés comme tous les scientifiques d’interpréter les données issues de l’observation et de l’expérience, et donc de passer par un langage de transposition, produisent nécessairement des configurations sémiotiques. Mais ces biologistes « asémiotiques » persistent à traiter ce langage comme une simple traduction métaphorique et approximative, en raison du caractère illusoire et trompeur qu’ils attribuent aux finalités du vivant. Autrement dit, ils ne prennent pas au sérieux leurs propres métaphores, et il serait pourtant nécessaire qu’ils le fassent, argumentent les biosémioticiens, pour redonner sa pleine consistance au langage de représentation et d’interprétation, c’est-à-dire à la couche sémiotique.
Si on prolonge le raisonnement, il ne serait pas possible d’interpréter correctement les résultats des recherches biologiques sans faire appel aux finalités, qui seules permettraient de donner une consistance sémiotique aux transpositions métaphoriques des biologistes. En d’autres termes, il faudrait d’abord admettre la pertinence des causes finales pour pouvoir prendre au sérieux l’interprétation sémiotique des faits biologiques : on peut tout de même au moins douter que la crédibilité de la sémiotique auprès des biologistes progresse sous l’effet de tels arguments !
Le terme et la notion qui cristallise la plupart des controverses est celle de « fonction ». Fonction est le terme piège par excellence, car c’est à la fois l’un de ceux dont la biologie ne peut pas se passer, et celui qui engage le plus facilement la projection de « buts » et de « finalités ». Darwin lui-même est impliqué dans cette discussion, car en remplaçant la cause finale lamarckienne par la notion de « fonction », sans préciser suffisamment ce qu’il entendait par « fonction », il a prêté le flanc à des critiques « anti-téléologiques ».
Si l’on dit par exemple que le cœur a pour fonction de pomper le sang, et si on considère que la fonction crée l’organe, alors la fonction est première, et on peut en induire que cette fonction est un but de la nature, celui qu’elle avait en faisant exister le cœur. Ce raisonnement peut être sémiotisé, comme dans l’extrait suivant, emprunté au même collectif d’auteurs (cf. supra) :
- Note de bas de page 15 :
-
Notre traduction. Kalevi Kull et al., « Theses on Biosemiotics », op. cit. , p. 168.
Les concepts de fonction et de sémiose (processus du signe) sont entrelacés. Les deux sont des concepts téléologiques au sens de déterminés par rapport à une fin — un contenu absent spécifiquement corrélé. Bien qu’on ne sache pas si ces deux propriétés des processus vivants (fonction et sémiose) sont exactement coextensifs, il est clair que, bien que les processus physiques irréversibles dans le temps se situent dans le monde physico-chimique prébiotique, les processus téléologiques qui sont spécialement organisés en ce qui concerne les fins ou les référents sont spécifiques aux processus du vivant. Si nous considérons une fonction comme un processus organisé autour d’une fin représentée implicitement, alors ces deux classes de phénomènes doivent être considérés comme coextensifs. Alternativement, la sémiose, l’activité des processus de signe peut être considérée seulement dans des conditions où il y a explicitement ou implicitement représentation d’un état final ou sous lequel une condition de satisfaction fonctionnelle peut être identifiée […], auquel cas la sémiose peut être définie en rapport avec la fonction antérieure.15
- Note de bas de page 16 :
-
On pourrait être tenté d’utiliser ici l’expression « lecture rétrospective ». Nous résistons à cette tentation en choisissant « lecture rétrojective » (on prend à rebours le processus), à laquelle s’opposera la « lecture projective » (on suit le cours du processus). Cela pour éviter i) d’impliquer trop vite et imprudemment la temporalité dans l’analyse du processus, ii) de fixer le point de vue autour d’un centre de référence qui serait celui de l’observateur et non celui de l’être vivant. (Cf. infra, 2.3.1).
L’assimilation entre une « fonction » et une « fin » implique de nouveau ici la conversion d’un élément absent (« un contenu absent spécifiquement corrélé », « une fin représentée implicitement ») en but, par la projection d’une temporalité irréversible sur la structure narrative. La téléologie est donc bien un effet du point de vue épistémique adopté, et tout le raisonnement s’effondre si on neutralise l’effet de sens temporel, notamment en adoptant la lecture — rétrojective16 et atemporelle — d’une sémiotique narrative modale. Nous y reviendrons.
Mais la question est toujours : est-ce de cette sémiotique-là que nous avons besoin pour traiter du vivant ? La transposition sémiotique repose sur la catégorie « présent / absent » (l’organe et ses propriétés sont présents, la fonction est absente) : cette catégorie fonde le signe : un representamen (présent) mis à la place d’un objet (absent) sous le contrôle d’un interprétant. Bien que l’absence de l’objet ne soit pas nécessaire à la construction du signe, elle déclenche ici, dans le cas du monde vivant, le raisonnement téléologique : si la fonction est « absente », c’est parce qu’elle ne peut pas recevoir de manifestations spécifiques directes, et qu’elle ne peut donc être saisie que lors d’une projection téléologique. Or un objet « absent » peut tout aussi bien être un objet reconstruit par catalyse : sous la pression des contraintes dynamiques d’un système, une case vide peut être remplie en raison même des contraintes qui pèsent sur le réseau de différences, et l’analyse (devenue alors, dans les termes de Hjelmslev, catalyse) peut identifier un jeu de différences locales (pour la case vide) qui obéit au réseau global des différences, et qui est actualisable en ce lieu particulier.
- Note de bas de page 17 :
-
Cet aspect de la théorie hjelmslevienne a été tout particulièrement mis en évidence par Antonino Bondi, dans « Hjelmslev et la “fonction sémiotique” : du modèle structural au modèle cognitif » (in Histoire Épistémologie Langage, tome 30, fascicule 2, 2008, « Découverte des langues à la Renaissance », pp. 199-212. )
Mais la notion de fonction peut être comprise tout autrement, et plus particulièrement d’un point de vue sémiotique. Si on suit la position de Hjelmslev dans sa construction d’une sémiotique en tant que stratification de niveaux, les strates de signification sont organisées en fonctions, chaque fonction étant une dépendance entre des fonctifs. Les fonctions sont des relations sélectionnées par les pratiques sémiotiques : la fonction sémiotique par excellence est une interdépendance entre un plan de l’expression et un plan du contenu, mais une interdépendance qui est toutefois instable, soumise à des déplacements et à des déformations, justement sous la pression des pratiques et des sélections qu’elles induisent17.
- Note de bas de page 18 :
-
Cela n’interdit pas de la convertir en « but », mais la construction de sa signification n’impose pas une telle conversion.
La sélection hjelmslevienne n’est évidemment pas la sélection darwinienne, mais la différence d’échelle et de substrat matériel ne doit pas masquer le fait qu’elle fonctionne, dans la perspective sémiotique, de la même manière : tout comme dans le domaine biologique, parmi la multitude, sinon l’infinité, des dépendances prévisibles ou imprévisibles, certaines sont sélectionnées, perdurent et se stabilisent (au moins provisoirement), et rendent possible la construction d’une signification. La fonction n’est pas un but, mais une dépendance sélectionnée qui devient, en raison de sa stabilité, structurante dans la stratification sémiotique18.
- Note de bas de page 19 :
-
Larry Wright, « Functions », The Philosophical Review, vol. 82, 2, 1973, pp. 139-168.
D’autres tentatives récentes, comme celle de Larry Wright, visant à neutraliser toute interprétation téléologique de la notion de « fonction », vont dans le même sens. Wright renverse le principe selon lequel la fonction crée l’organe, et affirme que l’organe n’existe que parce que la fonction résulte de l’organe19. En d’autres termes, la fonction n’est qu’un effet de la sélection : la fonction d’un organe est l’effet (et non le but) qui résulte du fait qu’il a été sélectionné ; en outre, la fonction a confirmé et stabilisé la sélection de l’organe. Une propriété biologique est sélectionnée dès lors qu’elle trouve une fonction durable, mais cette fonction est « inventée » par la sélection à partir de la propriété. Wright peut donc écrire en substance que l’ADN a été sélectionné et a perduré en raison du fait que cette sélection a eu pour effet de transmettre l’hérédité dans la réplication du vivant. Ce renversement de perspective, qualifié selon les auteurs d’« étiologique » ou de « rétrospectif », confirme la critique de l’illusion téléologique : ce qu’on prend pour une finalité n’est que l’effet d’une transformation obtenue après sélection, qu’on peut soit constater après-coup, soit prédire à partir des conditions initiales, et, dans tous les cas, considérer comme une condition requise pour la stabilisation du résultat. Une telle inversion, tout comme la conception de Hjelmslev, montre clairement la sensibilité des explications téléologiques aux effets de perspective, et donc aux choix méthodologiques qui président à l’interprétation : comme nous en faisions l’hypothèse en introduction, la question cruciale est bien celle de l’orientation épistémique de l’analyse.
Le raisonnement téléologique peut alors être compris comme la transformation non critique d’un effet de sens en but, sous une orientation épistémique projective et soumise à une temporalité irréversible. Reste à caractériser maintenant l’orientation épistémique que nous proposons.
2.2. Les « finalités » comme conditions narratives
- Note de bas de page 20 :
-
Aristote, Métaphysique, présentation et traduction par Marie-Paul Duminil et Annick Jaulin, Paris, Flammarion (coll. Garnier-Flammarion), 2008, Livre A.
Pour progresser en ce sens, nous pouvons revenir aux sources du problème, c’est-à-dire à la typologie des causes chez Aristote. La téléologie adopte les causes finales, parmi tous les types de causes possibles, comme explication principale ou directrice d’une situation, d’un événement ou de n’importe quel type de faits. Aristote distingue dans sa Métaphysique quatre types de causes : la cause matérielle, la cause formelle (l’eidos), la cause efficiente (la kinèsis) et la cause finale (le telos)20. Dans l’Éthique à Nicomaque, il superpose le télos et le Bien :
- Note de bas de page 21 :
-
Aristote, Éthique à Nicomaque, trad. Richard Bodéüs, Paris, GF Flammarion, 2004, livre I, 1094a.
Tout art et toute investigation, et pareillement toute action et tout choix tendent vers quelque bien, à ce qu’il semble. Aussi a-t-on déclaré avec raison que le Bien est ce à quoi toutes choses tendent.21
On peut alors supposer que le télos participe aux valeurs éthiques, celles qui guident ou orientent les processus de transformation. Si c’est bien le cas, elles doivent être actualisées et réalisables en tout point du processus concerné, et elles ne sont « finales » que parce qu’on suppose que leur réalisation complète et définitive ne peut advenir qu’après l’achèvement dudit processus.
Une autre lecture de la typologie aristotélicienne est donc possible si on part du principe qu’elle ne porte pas seulement sur les « causes » de l’existence des choses mais également sur leurs modes d’existence et sur les conditions qui rendent ces modes d’existence possibles et interprétables. Comme un mode d’existence ne peut être appréhendé qu’en contraste avec d’autres modes d’existence, et plus précisément à travers la transformation des modes d’existence, la typologie des causes aristotéliciennes peut être convertie, au moins comme hypothèse de travail, en une séquence narrative élémentaire, qui parcourt les modes virtuel, potentiel, actuel et réalisé.
La matière étant le substrat à transformer, elle passe du mode réel (elle impose sa résistance au début du processus) au mode potentiel (elle s’efface derrière la forme qui résulte du processus). Le mouvement (kinésis) est le processus de transformation lui-même, et il est donc en mode actuel de bout en bout. La forme (eidos) est le résultat manifestable du processus, et elle passe donc du mode potentiel (l’objectif fixé) au mode réalisé (le résultat final du processus). Le telos étant ce « au nom de quoi » un parcours est accompli en traversant toutes ses phases, il n’est jamais réalisé en tant que tel, et il reste indéfiniment virtuel : s’il s’actualisait ou se réalisait, ce ne serait pas une cause finale, mais une des causes efficientes. Ce parcours conduit alors d’un état initial (cause matérielle) jusqu’à un état axiologique visé (la cause finale, le télos), via une transformation et un mouvement (la cause efficiente, la kinesis) et un résultat reconnaissable (la cause formelle, l’eidos).
Sous la forme d’une séquence narrative, notre relecture de la conception aristotélicienne enchaînerait donc :
État initial (cause matérielle) > transformation (cause efficiente) > forme d’un résultat identifiable (cause eidétique) > état final (cause téléologique).
Il y aurait donc bien un mouvement prospectif dirigé vers le télos, m ais qui n’est pas celui de la construction de la signification — si ce n’est une signification purement spéculative, invérifiable, et dont l’actualisation peut être indéfiniment repoussée. La seule manifestation possible (mais indirecte) du télos résiderait dans l’intensité de la force d’engagement dans le parcours : la visée axiologique, en effet, s’incarne dans l’adhésion plus ou moins forte de l’actant à l’égard du processus. Mais pour construire la signification du télos proprement dit, dans notre relecture d’Aristote, il faut attendre que le processus soit accompli, que son télos soit effectivement réalisé, et que cette réalisation soit observable ou attestée, notamment en termes de véridiction. La signification ne pourrait être reconstruite et stabilisée qu’à partir de cette fin, si fin il y a. L’explication n’apparaît alors comme téléologique que si on fixe le point de vue au début du processus, ce point de vue où, par principe, on n’accède pas encore à la signification de ce dernier, du moins au sens où l’entend une sémiotique de la signification.
- Note de bas de page 22 :
-
Cette reformulation de la conception aristotélicienne s’accommoderait fort bien d’une lecture peircienne (à condition d’en neutraliser d’abord la pertinence narrative). Dans cette lecture, le « en raison de C » jouerait le rôle du fondement, qui guide le processus de la sémiose et notamment le dégagement d’un objet immédiat (équivalent de la cause formelle selon Aristote) à partir d’un objet dynamique (équivalent de la cause matérielle). Je laisse les spécialistes de l’approche peircienne apprécier si le « fondement » (le « en raison de C ») peut ou non être un but ou une perspective spéculative…
En l’absence de fin dernière manifestée, puisque l’accomplissement et la manifestation directe du télos sont indéfiniment repoussés, ce dernier ne peut en effet signifier en tant que cause que parce qu’il procure une médiation apparemment explicative entre un état initial matériel et un état final formel : la transformation entre A et B a lieu « en raison de C »22. Cette médiation serait alors une condition narrative, et non une finalité inaccessible. Le télos devient une médiation explicative dès qu’il est considéré comme une condition de la transformation. Cette condition ne peut être vérifiée que si la transformation a eu lieu, et cela implique, pour l’observateur, un raisonnement en deux temps : (1) une hypothèse, éventuellement une abduction, portant sur une condition réalisable et à réaliser au cours de la transformation, par conséquent de nature prospective, voire prédictive, et (2) une validation de l’hypothèse, par une construction rétrospective de la signification du parcours (d’où la valeur « explicative » de la condition).
L’orientation épistémique est alors renversée, car la validation de l’hypothèse ne peut être que portée par une lecture rétrojective et achronique. En outre, la conversion de la finalité en condition narrative impose un arrêt du processus, une fin (et non une finalité) à partir de laquelle la lecture rétrojective est possible.
2.3. Narrativités alternatives
Toutes les critiques adressées à la téléologie conduisent à reconfigurer les fonctionnements narratifs des processus décrits. En outre, à l’examen de ces critiques, on observe que l’étendue conceptuelle concernée augmente considérablement : les projections anthropomorphes, notamment, apparaissent soit comme des incitations à une téléologie qui remettrait en question la frontière non seulement entre les espèces, mais surtout entre culture et nature (les mobiles des conduites humaines sont transposés en lois générales de la nature), soit comme les conséquences d’une position téléologique (comme contagion métaphorique « prise au sérieux »). L’anthropomorphisme lui-même peut se dissimuler derrière un point de vue humain implicitement adopté et non discuté : il semble alors tout naturel de considérer que l’espèce humaine est le couronnement et le point ultime de l’évolution, alors que les lois de l’évolution disent tout le contraire. Les seules manières, pour les humains, d’être l’espèce ultime, seraient (1) de détruire toutes les autres espèces, et la planète Terre également, et/ou (2) de parvenir à un contrôle technologique de la réplication du vivant tel qu’il conduirait à une maîtrise de l’évolution. Mais même en ces cas extrêmes, on n’échappe pas aux principes de l’évolution et de la sélection : à des échelles de quelques centaines de milliers ou des millions d’années, ces catastrophes pourraient également être comprises comme l’un de ces accidents dont se nourrit le processus de la sélection, à condition bien entendu que les mondes vivants aient traversé ces accidents sans s’éteindre.
Dans toutes les critiques adressées à la téléologie, des alternatives narratives et modales sont impliquées. On se demande pour commencer s’il y a, ou non, un « actant » directeur ou régulateur. S’il y en a un, on s’interroge ensuite : est-il externe ou transcendant (Platon, ou le dessein intelligent) ? Est-il interne ou immanent (Aristote, Spinoza) ? Est-il anthropomorphe ou est-il seulement une force, une énergie impliquée dans le système (Leibnitz, ou la cybernétique) ? Est-il exprimé et observable ? Est-il caché ? S’il n’y a pas d’actant à proprement parler, y a-t-il au moins des dominantes ou des inflexions modales qui permettraient de construire la signification des processus décrits ? On a vu le rôle du possible, du nécessaire et du contingent, de l’imprévisible et de l’aléa. On pourrait évoquer aussi (cf. infra) la construction d’une position subjectale dans l’Umwelt, chez von Uexküll, comme premier point d’appui pour une construction du sens des interactions naturelles.
Mais on remarque surtout, dans les différents débats, le rôle de l’observateur : selon qu’il se positionne prospectivement ou rétrospectivement, il postulera des projets, des buts, des finalités, ou des résultats, des destins et des conditions nécessaires, des effets sélectionnés ; selon qu’il sera lui-même présent en tant qu’être vivant, ou limité à un rôle cognitif, ou enfin réduit à une pure position de référence, il effectuera des projections anthropomorphes, il procédera à des évaluations modales épistémiques, il variera son point de vue, ou il ne sera qu’un centre d’orientation et de mémorisation des interactions.
2.3.1. Lecture à rebours et modalisante
- Note de bas de page 23 :
-
Claude Bremond, « La logique des possibles narratifs », Communications, 8, Recherches sémiologiques : l’analyse structurale du récit, 1966, pp. 60-76.
La sémiotique narrative s’est elle-même trouvée devant de telles alternatives, depuis les années fondatrices au milieu du XXe siècle. Les solutions retenues sont principalement de nature méthodologique : face à quelque objet d’analyse que ce soit, l’analyse narrative suppose un ordre de transformation qui dispose des avant (A) et des après (B), en bref une succession signifiante. Cette succession n’est pas nécessairement celle de l’objet à analyser car elle est de nature positionnelle et non temporelle ; mais l’analyste doit se demander dans quel sens il doit construire la signification de cette succession : de A vers B, ou l’inverse ? En lecture projective (vers B, vers la fin ultime), ou en lecture rétrojective (vers A, vers le point d’origine du processus) ? La disposition matérielle dominante des objets d’analyse (par exemple la manière de raconter les histoires) invite tout naturellement à adopter la lecture projective, une sorte de « sémiotique par défaut », de bon sens, qui commence par le commencement (A) et finit par la fin (B). C’était le principe retenu par Claude Bremond dans sa méthode des possibles narratifs reposant sur l’identification préalable d’un point d’origine, puis de points de bifurcation ultérieurs23.
Bien entendu, en lecture projective (comme dans le raisonnement téléologique), il faut postuler et disposer d’un « en raison de C » (on passe de A vers B en raison de C) susceptible d’expliquer les choix parmi tous les possibles. Dans ce type de méthode projective, les deux situations qui se présentent sont insatisfaisantes : ou bien « en raison de C » change à chaque bifurcation, voire reste entièrement aléatoire, et la reproductibilité comme la prédictibilité sont nulles, ou bien « en raison de C » est toujours le même, et c’est alors un « actant caché », le Deus ex machina, ou a minima une intentionnalité diffuse mais directrice qui explique cette régularité : même si nous ne pouvons pas savoir vers quoi se dirige le processus, lui, il le « sait ».
En revanche, la lecture rétrojective, de B vers A, neutralise la temporalité en prenant à rebours l’ordre de succession des phases du processus, et ce faisant, elle va identifier des opérations de transition par modalisation. En effet, en partant de la situation finale, elle rencontre elle aussi des points de bifurcation, mais qui apparaissent chacun comme contrôlé par une modalité (un devoir-faire, un vouloir-faire, un savoir-faire ou un pouvoir-faire). En d’autres termes, ce qui apparaît comme une quantification et une sélection aléatoire des possibles dans le sens projectif sera traité comme modalisation du processus dans le sens rétrojectif. C’est très précisément ce qui se passe quand nous convertissons (cf. supra) une cause finale en une condition nécessaire (une nécessité à la place d’une finalité), ou bien un but de la sélection en un effet sélectionné : nous inversons l’ordre de lecture. En lecture projective, « en raison de C » est un but, un objectif ou une finalité ; en lecture rétrojective, « en raison de C » est une modalisation du processus sémiotique.
Nous avons choisi de qualifier l’orientation méthodologique de projective ou rétrojective, plutôt que « prospective » et « rétrospective » pour trois raisons :
i) tout d’abord pour ne pas subjectiver le choix de manière intempestive et a priori ; la prospection et la rétrospection se réfèrent en effet non pas à la structure d’un processus, mais à la position subjective et contingente d’un observateur ; la projection et la rétrojection font également appel à deux points de vue différents, mais qui sont supposés déjà inscrits (cf. supra, phase A et phase B) dans la structure du processus lui-même ;
ii) ensuite pour éviter de mêler le temps à cette affaire : la transformation narrative est une structure dynamique mais achronique, dont l’interprétation, soit dans un sens soit dans l’autre, ne consiste pas à suivre ou à remonter le temps, mais à prendre position par rapport à la structure elle-même. Certes, l’expérience d’une transformation narrative est nécessairement temporelle (mais aussi spatiale) : ce n’est pas une raison suffisante pour que l’interprétation narrative soit elle-même sans autre précaution confondue avec une expérience temporelle ;
iii) enfin, pour bien distinguer les opérations portant sur la structure narrative et les opérations portant sur le temps : une lecture rétrojective (de B vers A) n’interdit pas de prédire ou d’anticiper B (temporellement) à partir de A. C’est précisément ce que nous avons déjà laissé entendre, en distinguant la prévision, la prédiction et la projection d’une finalité : l’hypothèse projette temporellement une prévision mais ne peut être validée et assumée (c’est-à-dire interprétable) qu’en lecture rétrojective.
Ce sera donc la première condition pour concevoir une sémiotique du vivant qui ne soit pas tentée par la téléologie : la lecture rétrojective et modalisante, pour pouvoir construire la signification du processus décrit.
2.3.2. Une étude de cas : lecture projective et lecture rétrojective du code génétique
La génétique est probablement le domaine de la biologie où la question de l’autonomie du vivant par rapport aux systèmes physico-chimiques se pose avec le plus d’acuité, en raison justement de l’étroite intrication, à cette échelle d’analyse, entre ces derniers et l’émergence de la vie. C’est pourquoi les biosémioticiens se focalisent notamment sur le code génétique pour justifier l’émergence d’une dimension sémiotique en même temps que celle du vivant.
Howard Pattee, physicien et biologiste, a élaboré une solution simple et lumineuse pour rendre compte du prétendu « saut ontologique » entre les systèmes physiques et les systèmes vivants. Il commence par circonscrire le problème à traiter en caractérisant les systèmes biologiques, en contraste avec les systèmes physiques, par leur capacité d’auto- réplication et de conservation matérielle de la mémoire de ces réplications. Il faut donc comprendre pourquoi certaines structures physico-chimiques donnent lieu à des formes qui sont en mesure de se répliquer elles-mêmes de manière fiable, quel que soit le nombre de réplications : tout se passe comme si ces systèmes vivants étaient dotés d’une mémoire, comme si les formes qu’ils prennent pouvaient être ensuite indéfiniment « héritées » par d’autres systèmes vivants.
- Note de bas de page 24 :
-
Notre traduction. Howard H. Pattee, « La physique des symboles : combler la coupure épistémique », Biosystèmes, vol. 60, 2001, pp. 5-21.
Mais au lieu d’attribuer ces différences entre systèmes physiques et biologiques à un changement de domaine ontologique, il fait l’hypothèse d’une « coupure épistémique » entre les lois dynamiques du monde physique en général, et les jeux de contraintes qui sont spécifiques aux systèmes vivants, et qui expliqueraient leurs capacités d’héritage. Tout comme pour la cybernétique, il s’agit de « systèmes primaires » et de « systèmes secondaires », réunis dans une boucle de rétroaction, mais donc la séparation est de nature épistémique, et non pas ontologique. Pattee n’introduit aucune finalité dans les systèmes secondaires de contrôle, et, mieux encore, il impute l’apparente finalisation du vivant au couplage entre ces systèmes de contrôle et les conditions propres à l’observation. Ce raisonnement, imaginé et transposé à partir de celui qui prévaut en mécanique quantique, est alors résumé ainsi : « La mesure transforme des états physiques en symboles, et les contrôles mémorisés transforment les symboles en états physiques »24.
La coupure épistémique n’est donc que dans l’acte même d’observation, et elle se résout en une série où le vivant occupe une position médiane entre deux états des systèmes physiques :
[systèmes physiques1 > /mesure>symboles>mémoire/ > systèmes physiques2]
Il n’y a de saut entre le physique et le vivant que si nous cherchons à observer et à connaître ce dernier. Mais dans ce cas la coupure épistémique est inévitable, comme en mécanique quantique, en raison des paramètres spécifiques des systèmes physiques sous-jacents au vivant (vitesse de l’évolution de l’énergie, présence ou pas d’alternatives à sélectionner, etc.). L’apparente autonomie des symboles (la partie sémiotique des systèmes) est une pure illusion, écrit Pattee, et pourtant cette illusion ne peut être réduite, puisqu’elle participe à la production des états physiques en tant qu’objets connaissables (cf. supra).
Ontologiquement, les deux types de systèmes sont inséparables. Scientifiquement, dès qu’on les observe, on interagit avec eux, on induit une sorte d’autonomie méthodologique du vivant, cette séparation entre lois physiques et contraintes symboliques étant compensée par le cycle des rétroactions, qui réparent en quelque sorte la séparation induite par l’observation. Par conséquent, une fois encore, l’étroite interaction de l’observateur avec les mondes vivants est décisive, non seulement parce qu’elle rend leur connaissance possible, mais aussi parce qu’elle est le point critique où se décident les principales orientations d’une sémiotique du vivant.
Dans un article intitulé « Sémiotique et biologie », Denis Bertrand et Bruno Canque évoquent eux aussi, à propos de la génétique, le passage du physico-chimique au biologique :
- Note de bas de page 25 :
-
Denis Bertrand et Bruno Canque, « Sémiotique et biologie », in Amir Biglari (éd.), Sémiotique en interface, Paris, Kimè, 2017, pp. 600-601.
(…) le passage du physique au biologique s’opérait par l’établissement de correspondances terme à terme entre des bio-polymères de natures chimiques distinctes (telles qu’observées entre les chaînes d’acides nucléiques et les chaînes d’acides aminés formant la séquence des protéines) comme étant la condition initiale, autrement dit le préalable à la fois nécessaire et suffisant, de ce passage par la mise en acte de l’information : les correspondances terme à terme créent les conditions de son transfert et de son actualisation. Nous avions alors considéré que l’établissement de cette correspondance initiale, au terme d’un processus de réification et de linéarisation, pouvait être vu comme le fondement du concept d’information biologique et l’origine des processus évolutifs. D’un point de vue dynamique, cette correspondance permet également d’assurer à la fois le stockage, le transfert et l’exécution de ladite information. Selon cette perspective, nous en avions conclu qu’il n’y a pas d’opposition ni même de discontinuité entre le monde vivant et le monde physique. L’émergence des phénomènes biologiques devait plutôt être considérée comme l’un des prolongements possibles des processus physico-chimiques. Cette approche présente l’intérêt d’insérer la problématique de l’origine de la vie dans une dynamique évolutive qu’on peut qualifier d’évolution anté-historique, fidèle au gradualisme darwinien, ayant comme principal avantage de rendre caduque la recherche d’un temps zéro du vivant, d’un hypothétique « big bang » biologique qu’en d’autres temps on eût appelé Création.25
Les relations entre « les chaînes d’acides nucléiques » (qui constituent les ADN et les ARN) et les « chaînes d’acides aminés » (qui participent à la construction des peptides et des protéines) sont prudemment caractérisées par Bertrand et Canque comme des correspondances. Les généticiens sont souvent plus audacieux, notamment lorsqu’ils évoquent les « fonctions » de l’ARN : messager, transfert, réplication, traduction ou inhibition de la traduction. Concrètement, et d’un point de vue biologique, il s’agit en fait d’une série d’ARN (et donc d’acides nucléiques) dont la constitution est appropriée, en raison de la sélection opérée par l’évolution du vivant, aux rôles qu’ils jouent, et il y a donc des structures biologiques spécifiques de l’ARN messager, de l’ARN de transfert, une autre de l’ANR catalytique, etc.
On voit bien ici comment une dérive de l’approche sémiotique pourrait s’insinuer dans ce que Bertrand et Canque ne désignent que comme des correspondances : d’autres, bien plus nombreux, projettent des fonctions (des actions thématiques) sur ces correspondances, et ces fonctions conduisent à inférer des actants thématiques, voire un « actant caché » (un « code-maker ») dans la machine biologique. Une micro-narrativité émerge alors d’une analyse phénoménologique.
Mais on voit tout aussi bien pourquoi cette phénoménologie sémiotique est intempestive : faute d’explications biologiques suffisantes, et de résultats expérimentaux probants, le phéno-biologiste et le biosémioticien imaginent une frontière ontologique qu’un actant caché va aider à franchir, puis prêtent à ce dernier des « rôles actantiels » thématiques (communiquer, transmettre, créer, traduire, etc.). Ce point est tout particulièrement évident dans l’article déjà cité de Kull et al. (supra 2.1.2).
Dans ce même article, les auteurs prennent ensuite le cas de l’hémoglobine pour montrer que la « fonction » (le transport de l’oxygène) reposerait sur la relation entre un « absent » pourtant nécessaire, l’oxygène, et une région de la molécule qui est appropriée pour le capter et le transporter. Mais ils n’échappent pas à l’extrapolation métaphorique, puisque ce « site » d’association convertit selon eux la molécule d’hémoglobine en « signe-véhicule ».
Si on considère maintenant le rapport entre le génotype (l’ensemble structuré des ADN et ARN) et la production des composants organiques, les protéines, la médiation est assurée principalement par des molécules catalytiques, les enzymes (une partie des protéines fabriquées à partir des acides aminés, produits eux-mêmes par l’intermédiaire des ARN), et des ribozymes, directement constituées d’ARN (cf. supra, l’ARN catalytique). On pourrait donc supposer que cette médiation biochimique suffit à maintenir un lien de ferme détermination, garant de la reproductibilité des résultats. Mais les choses se compliquent. Sans entrer dans le détail de cette complexité, on peut se contenter de dire comme la plupart des généticiens que, du moins dès qu’on les observe, les correspondances n’apparaissent pas linéaires.
La clé du problème se situe dans l’expression biochimique du gène, c’est-à-dire principalement dans les modalités de fonctionnement de l’ARN dit « médiateur » : ce fonctionnement peut être affecté de multiples manières (par les interactions avec l’environnement, par des interactions multilatérales entre protéines et enzymes, ou même par des inhibitions de telle ou telle partie du gène, selon la forme des enroulements des ADN) ; ces modifications peuvent être durables, notamment grâce à des « transcriptions inverses » de l’ARN vers l’ADN : on peut ainsi hériter des empreintes biologiques issues d’interactions antérieures, notamment traumatiques, à travers plusieurs générations.
- Note de bas de page 26 :
-
« Qu’est-ce qu’un gène ? », La recherche, 348, 2001. Cet article a fait l’objet d’une analyse antérieure, et dans une autre perspective, dans J. Fontanille, « L’hétérogénéité des corpus : approches sémiotiques. Dix-huit définitions du gène », in Bernard Castagna (éd.), Du traitement du discours dans les recherches en communication, Les Cahiers de jérico, université de Tours, 4, 2004.
Afin d’entrer plus précisément dans le problème, nous pouvons nous référer à un dossier de la revue La Recherche qui, présentant une série de dix-huit définitions du gène, mit en évidence de profonds désaccords chez les biologistes26. Ces désaccords prennent leur origine dans la superposition problématique, voire impossible, entre deux types de définitions du gène, en tant que séquence moléculaire matérielle, et en tant qu’unité d’information.
La définition en tant que séquence moléculaire parcourt le processus des apparitions, disparitions, transpositions et mélanges des différents types de constituants, tout au long d’une séquence d’opérations de production :
a) la régulation de la région d’ADN dite « ADN promoteur » ;
b) la transcription de l’ADN promoteur par l’« ARN transcripteur » (une enzyme polymérase), qui peut produire un intermédiaire, l’« ARN messager » ; certaines régions de l’ADN promoteur ne sont pas transcrites (les segments dits « introns ») ; le début et la fin de la séquence transcrite sont indiqués par deux nucléotides, des « codons », respectivement le « codon start » et le « codon stop » ;
c) la synthèse : l’ARN transcrit et un acide aminé sont assemblés grâce à une molécule intermédiaire, un ARN dit « ARN de transfert », et on peut alors dire que l’ARN transcrit a synthétisé une protéine ;
d) la maturation : la protéine synthétisée subit de nombreuses modifications sous l’effet de son environnement chimique.
La définition en tant qu’unité d’information est elle aussi soumise à de grandes variations, mais de nature cognitive et épistémique : a) pour la synthèse de protéines, le gène « code » ; b) il est une « instruction » pour un programme génétique ; c) il « détermine » l’apparition d’un caractère phénotypique ; d) la diffusion d’un gène est « statistiquement corrélée » à la diffusion d’un phénotype ; e) le gène « conditionne » deux propriétés essentielles et indissociables du vivant : sa reproductibilité et sa variabilité.
Bien que les résultats expérimentaux soient dits scientifiquement « significatifs », les divergences entre les différentes interprétations de ces résultats brouillent la compréhension de leur « signification ». A la différence de l’interprétation en tant que séquence moléculaire, l’interprétation en termes de processus informatif s’empare du gène comme un tout, comme un actant cognitif responsable d’opérations dont les produits sont des observables dans le monde vivant. Cet actant est doté en effet de véritables (et peu vraisemblables) propriétés cognitives : marquage, instruction, programmation, conditionnement. Ces opérations se distribuent, selon les interprètes, tous biologistes, sur une gamme de valeurs épistémiques, depuis la causalité jusqu’à une simple corrélation statistique, depuis le « certain » (le gène code, fournit des instructions pour un programme) jusqu’au seulement « probable » (le gène conditionne, est corrélé statistiquement).
L’interprétation selon la corrélation statistique, qui est aujourd’hui dominante, induit un saut qualitatif irréversible : elle interdit notamment de rabattre la protéine sur le gène ; on peut « remonter » de la protéine produite jusqu’au gène qui en est l’origine, mais non l’inverse. L’une des principales difficultés tient à la quantification de chaque phase du processus. On a affaire à une syntaxe d’opérations, et la propriété critique de cette syntaxe est son aspectualité, et plus précisément la quantification qui va s’appliquer à chacune des phases aspectuelles. Au fur et à mesure qu’on avance dans la chaîne de production des figures matérielles, la quantification des opérations, par un effet cumulatif, multiplie les alternatives et les alternatives d’alternatives, interdisant de ce fait la rétroaction causale, et obligeant à un saut épistémique.
Les zones critiques aspectuelles sont les suivantes :
a) Il y a toujours sur l’ADN plusieurs régions de fixation possibles pour l’ARN polymérase : le « choix » de l’ARN promoteur est donc multiple.
b) Pour le même promoteur ADN, il y a toujours plusieurs bornes ou sites de début et de fin : le choix de la séquence codante précise est donc lui aussi multiple (plusieurs codons « start », plusieurs codons « stop »).
c) Lors de la transcription, le phénomène de l’épissage des brins d’ADN multiplie les possibilités de production de protéines : au cours du processus de duplication, l’ARN polymérase qui suit la séquence ADN de référence peut dupliquer les deux brins de l’hélice à la fois, un seul d’entre eux, ou, le plus souvent, sauter de l’un à l’autre, selon un rythme et une fréquence eux-mêmes multiples.
d) A partir d’un même ARN messager, on peut obtenir une multitude d’ARN transcrits, selon les interactions en cours.
e) Les conditions de maturation des protéines synthétisées peuvent enfin produire une multitude de variétés.
Globalement, la lecture projective du processus (du gène vers la protéine) est donc indéterminée, et l’observateur doit se satisfaire de probabilités, alors même qu’on dote le système génétique d’un actant cognitif responsable d’opérations précises ! En chacune des phases aspectuelles, chacun des facteurs de pluralisation implique un tri (un micro-événement de sélection) : il y a plusieurs solutions possibles, une seule est réalisée à la fois, et une instance de contrôle est à construire. Aussi longtemps que le « triage » ne peut pas être rapporté à des opérations chimiques spécifiques, une hypothétique instance cognitive de contrôle en prend la place. Mais les noms thématiques (message, transfert, traduction, etc.) des actes de cet actant cognitif masquent sans les résoudre les multitudes d’alternatives cumulées.
L’interprétation en termes d’information génétique, avec les problèmes épistémiques associés, n’apparaît qu’en lecture « projective » du processus de production, du gène vers la protéine, quand la relation de causalité n’est plus inférentielle (Si A est réel, alors B est actuel), mais sélective (Si A1, A2, A3, A4, etc., sont potentiels, alors B est actuel, en raison de C). Le problème, comme nous l’avons déjà signalé, c’est « en raison de C » : on parle d’autant plus d’information et de code qu’on maîtrise moins la relation matérielle entre le dispositif de référence et ce qu’il semble produire. Dans le sens projectif, la quantification et les opérations de sélection provoquent donc une conversion sémiotique qui fait appel à un actant masqué (pour jouer le rôle de C). Quand on ne sait pas clairement pourquoi tel gène produit telle protéine, on affirme que le gène « code » cette protéine et pas une autre. Et comme ce n’est pas une explication décisive, on ajoute que tout cela est contrôlé par un « code maker ». Mais c’est alors ce qu’on pourrait appeler une « sémiotique par défaut d’explication », et non une sémiotique qui participerait directement à l’explication biologique. Donnons la parole à l’inventeur du « code maker », Marcelo Barbieri :
- Note de bas de page 27 :
-
Marcello Barbieri, « Biosemiotics : A new understanding of life », Naturwissenschaften, 95, 7, 2008, pp. 25-26.
The components of sign do not come into existence by themselves ; they are produced by the same agent named a codemaker. […] Proteins, essential to a variety of functions in living organisms, are considered “manufactured” molecules.27
L’actant masqué est ici démasqué : si les correspondances physiques ne sont pas suffisamment déterministes, il faut supposer un « code », une convention qui serait d’une autre nature que les règles biologiques, dont il faut alors imputer la responsabilité à une instance qui n’est pas physico-chimique. Si on pose d’abord une frontière (une discontinuité ontologique) entre le monde physique et le vivant, il faut inventer un actant cognitif pour la franchir, mais un actant cognitif qui n’est pas l’observateur, et qui est dans les systèmes eux-mêmes. Et c’est très précisément dans cette zone de quantification aspectuelle problématique (entre ADN, ARN et protéines), que les fonctions de tri sont imputées à un code, dont la production est elle-même imputée à un codemaker. La solution de Barbieri illustre parfaitement les difficultés épistémologiques et méthodologiques inhérentes à une « sémiotique par défaut » : le sémioticien, c’est celui qui confie à un « actant caché » l’explication de ce qu’on ne comprend pas (encore), ou de ce qui obéit à des règles qu’on peine (pour le moment) à expliciter.
En revanche, la lecture rétrojective, de la protéine vers le gène, est de même nature que celle de la construction de la signification suivant la logique narrative, selon laquelle, en partant de la situation finale, chaque point de bifurcation apparaît rétrospectivement comme contrôlé par une modalité (en l’occurrence, un devoir-faire, un savoir-faire ou un pouvoir-faire). En d’autres termes, ce qui apparaît comme une quantification épistémique des possibles dans le sens projectif sera traité comme modalisation du processus dans le sens rétrojectif.
Dans cette conversion de l’unité d’information en « structure matérielle », en lecture rétrojective, la quantification s’inverse (ou réduit un certain nombre d’entités produite à une seule entité productrice) et la question du tri ou du choix ne se pose plus : il y a plusieurs protéines possibles issues d’un même gène, mais en partant d’une ou plusieurs protéines données, on peut remonter au gène unique ou au groupe de gènes qui lui ou leur correspond.
La différence entre lecture projective et lecture rétrojective est relative à un point de vue de l’observateur, et plus généralement à l’orientation épistémique de l’interprétation, et cette différence d’orientation induit deux manières différentes de construire la signification. Les deux types de sémiotisation sont : i) dans le sens projectif, une sémiotique cognitive qui associe des indices et des expressions à des contenus d’information, et qui doit expliquer la raison des tris effectués ; ii) dans le sens rétrojectif, une sémiotique pratique qui reconstitue et modalise des modus operandi, ce qu’on pourrait appeler aujourd’hui une « sémiotique des empreintes d’interactions », associée à une sémiotique modale. Cette biosémiotique-là focaliserait sur les modalités de l’inscription sur un support matériel, sur les interactions avec ce support et sur la syntaxe modale qui les régule. La première ouvre la porte à des résultats non vérifiables et non reproductibles, notamment parce qu’elle implique un « actant caché » supposé faire des choix en fonction de buts à poursuivre. En revanche, la seconde, qui reste en tout point sensible aux contraintes pratiques et matérielles, ne provoque pas de rupture majeure avec le principe de falsification et de reproduction des résultats qui s’impose à la biologie.
3. Sémiotique des interactions et régimes de signification
- Note de bas de page 28 :
-
E. Landowski, Les interactions risquées, Nouveaux Actes Sémiotiques, 101-103, 2005, ainsi que Passions sans nom, Paris, PUF, 2004.
La seconde condition que nous posons, pour la sémiotique du vivant que nous envisageons, sera de nature actantielle, et elle ouvre sur une narrativité alternative radicale, dont la différence ne repose pas seulement sur un point de vue méthodologique, mais sur une conception spécifique des conditions à mettre en place pour la construction de la signification. La socio-sémiotique d’Eric Landowski offre à cet égard déjà une alternative, ou plutôt un principe général englobant, qui doit ici retenir notre attention28 : si nous voulons accéder à des organisations signifiantes sans projection anthropomorphe, nous devons à l’évidence ne pas nous précipiter sur les structures actantielles greimassiennes, que leur inventeur qualifiait lui-même d’« anthropomorphes ». Autrement dit, les concepts de destinateur, destinataire, sujet, objet, etc. ne sont pas encore disponibles à ces échelles d’analyse. Pas plus, d’ailleurs, que les opérations de la jonction, qui participent elles aussi d’une syntaxe anthropomorphe.
3.1. Des interactants non spécialisés impliqués dans des régimes sémiotiques
Eric Landowski pose, au départ de toute possibilité de signification, les interactions. Au cours des interactions, deux événements décisifs se produisent. D’un côté, des rôles se dessinent, dans la mesure où les interactions induisent des relations symétriques ou dissymétriques, et notamment la dissymétrie entre soi et autrui, et aussi des esquisses de sujets et des ébauches d’objets, mais dans tous les cas ces relations restent réversibles et ces rôles, interchangeables. De l’autre côté, des régimes de sens prennent forme (des plans, des programmes, des aléas, des ajustements, etc.). Sans le dire aussi explicitement, Uexküll procède également ainsi pour décrire le fonctionnement des Umwelten, qui sont globalement et avant toute chose des espaces d’interactions. Dans une sémiotique des interactions, il n’y a pas encore nécessairement de communication, d’échange, ou de prédation : tout cela se construit progressivement, à partir des régimes de signification qui émergent par schématisation des interactions elles-mêmes. Ce qu’on recherche donc en premier lieu, ce sont des régimes de signification, plus ou moins élaborés, hiérarchisés, et mis en relation entre eux, et ce ne sont pas des signes ou des systèmes de signes. Les Umwelten de von Uexküll sont, comme nous le montrerons tout à l’heure, de tels régimes de signification, clairement plus pertinents, sophistiqués et explicatifs que les « signes » qu’il manipule aussi par ailleurs.
- Note de bas de page 29 :
-
J. Fontanille, Sémiotique du visible. Des mondes de lumière, Paris, PUF, 1995 ; id., Sémiotique du discours, Limoges, PULIM, 2003.
Cette question posée à la sémiotique narrative n’est pourtant pas neuve. Dans Sémiotique du visible. Des mondes de lumière, et dans Sémiotique du discours, nous proposions de compléter la structure actantielle narrative (destinateur, destinataire, sujet et objet) élaborée par Greimas, avec la série source / contrôle / cible, pour traiter notamment de phénomènes propres au monde naturel et qui, bien que relevant d’une narrativité profonde, ne pouvaient être saisis sans un biais fort discutable par la structure actantielle anthropomorphe29. Nous distinguions alors les actants transformationnels (ceux de la narrativité greimassienne) et les actants positionnels (la série : source / contrôle / cible). Ces derniers n’impliquent en particulier aucune intentionnalité narrative, et n’exigent que trois prémisses : (1) une position de référence, (2) une orientation qui en découle, (3) une régulation (le contrôle). Cette proposition n’a guère eu de succès, peut-être parce qu’elle était trop simple, et probablement parce qu’il y avait alors peu d’occasions ou de matière pour la mettre en œuvre. Or, si la biosémiotique se (re)construit comme une sémiotique des interactions, les interactions au sein des mondes vivants doivent nécessairement être orientées et régulées : ces deux clauses sont au fondement même, par exemple, de l’éthosémiotique de von Uexküll.
Pour en faire un nouvel usage aujourd’hui, peut-être faudrait-il préciser que « source » et « cible » sont des dénominations dont le caractère figuratif n’a aucune incidence pertinente : seule compte en l’occurrence l’orientation entre les deux instances, et le fait que l’une des deux, provisoirement sélectionnée comme centre de référence de cette orientation, vise l’autre. La visée confirme l’orientation, mais ajoute, ce qui est essentiel, un mouvement sensible. Par conséquent, la structure actantielle minimale dont nous avons besoin est une relation sensible et orientée entre deux instances, qui est régulée par une tierce instance. Par exemple : une tique, un mammifère et un Umwelt.
- Note de bas de page 30 :
-
E. Landowski, « Avoir prise, donner prise », Actes Sémiotiques, 112, 2009.
Dans un article très dense consacré à nos relations avec les objets, Eric Landowski a lui aussi, quelques années plus tard, et indépendamment de la proposition précédente, élaboré une solution pour traiter les structures actantielles minimales qui émergent dans les interactions30. Son objectif était également de rendre compte d’interactions non intentionnelles, et d’éviter la projection précipitée des structures actantielles anthropomorphes sur des phénomènes qui ne le justifiaient pas. Néanmoins, la proposition doit être reçue avec précaution, dans la mesure où la moitié seulement de l’anthroporphisme est éliminée. Il précise notamment au début de l’article :
(…) si la même dimension sensible imprègne toutes nos constructions de sens, et si ce dont il s’agit de rendre compte c’est de l’expérience concrète du sens dans nos rapports au monde en général, il ne peut y avoir qu’une seule sémiotique dont la vocation est d’englober l’ensemble de nos relations à « l’Autre », qu’il s’agisse de nos « semblables » ou de n’importe quelle grandeur « inanimée » peuplant notre environnement.
L’« autre » peut donc être n’importe quel type d’existant, mais le Soi (nous, nos) est bien humain (trop humain ?). Toutefois, ce demi-anthropomorphisme n’a pas d’incidence repérable dans le modèle général proposé. Il est d’abord formulé à partir de la notion de « prise » : la relation entre soi et l’autre est dissymétrique, donc orientée, car pour soi il est question d’« avoir prise » sur l’autre, et pour l’autre, de « donner prise » à soi (quitte cependant à devoir pour cela donner soi-même, stratégiquement, prise à cet autre). Comme cette relation de prise est d’emblée définie comme d’ordre sensible, elle n’est pas si éloignée de la notion antérieure de visée. Cette première modélisation, projetée sur une interaction, et plus précisément sur les opérations qui constituent les interactions, donne lieu à une série actantielle : operator / operans / operandum.
On reconnaît aisément dans l’operator et l’operandum, respectivement l’équivalent de la source et de la cible de l’opération. Entre les deux, l’operans est une instance qui peut se manifester comme outil, instrument, moyen, milieu, tout ce qui en bref constitue une médiation (le plus souvent modale, de l’ordre du savoir ou pouvoir faire) entre l’operator et l’operandum, comme le précise Landowski lui-même :
- Note de bas de page 31 :
-
« Avoir prise, donner prise », art. cit., 2e partie, II.1, in fine.
Ce qui en ressort sur un plan général, c’est que toute relation entre un operator et son operandum étant nécessairement relayée par la médiation de quelque operans, chacune de ces médiations se trouve elle-même relayée par quelque autre médiation.31
Nous ne sommes pas donc si loin également de l’instance de contrôle qui, par définition est en situation de médiation en même temps que de régulation. A ce point de ce petit exercice de mémoire, il importe peu de choisir entre l’une ou l’autre formulation. En revanche, les points de consensus offrent un point de départ possible pour doter la sémiotique des interactions dans les mondes vivants d’une dimension syntagmatique minimale. Elle sera définie comme
i) l’établissement d’une relation orientée entre au moins deux interactants,
ii) la sensibilisation de la relation et des deux interactants (visée ou prise, peut-être même les deux),
iii) une médiation dans cette relation elle-même, comprenant notamment toutes les propriétés modales, thématiques, voire affectives et somatiques qui caractérisent et régulent le devenir commun des deux interactants, et enfin
iv) une réversibilité, synchrone ou asynchrone, de la relation sensible en question. Alors peuvent se déployer des régimes de signification, dont les variables sont au moins la présence ou l’absence de l’une de ces quatre caractéristiques, mais aussi et surtout les différentes propriétés de l’instance de contrôle et de médiation (en l’occurrence l’Umwelt lui-même).
Pour illustrer l’antécédence des interactions et des régimes de signification sur les rôles actantiels spécialisés, on peut faire appel à une critique que von Uexküll s’adresse à lui-même, à propos de sa description de l’Umwelt de la tique — dont il dit qu’elle « guette sa proie » :
- Note de bas de page 32 :
-
J. von Uexküll, Milieu animal et milieu humain, trad. Charles Martin-Freville, Paris, Payot, 2010, p. 97.
Mais déjà pour décrire la vie de la tique, nous avions dit qu’elle guettait sa proie. A travers cette expression, même si c’est involontaire, nous avons déjà laissé les petits tracas de la vie quotidienne des humains s’immiscer dans la vie de la tique qui est uniquement commandée par le plan de la nature.32
Uexküll reconnaît qu’il anticipe un stade d’élaboration de l’Umwelt qui n’est pas celui de la tique. Il pourrait être animal, et pas seulement humain, mais bien au-delà de celui de la tique. La tique ne guette rien, elle est seulement immobile et non stimulée ; si le stimulus advient, elle se relâche et elle tombe, ce qui déclenche une interaction strictement programmée avec un mammifère qui n’est pas à proprement parler une proie, mais seulement le « site » (la cible et l’operandum) où se produira l’interaction principale, celle qui procure le sang nécessaire à la ponte des œufs. Sans aucun anthropomorphisme, on pourra alors repérer des phases contrôlées par le pouvoir-faire ou le pouvoir-être (pouvoir ne pas tomber / pouvoir tomber, et se laisser tomber sur le mammifère-hôte) d’autres par le savoir-faire (chercher et trouver une zone de poils moins dense ; perforer la peau et tester la chaleur du liquide aspiré), etc. On voit bien alors que si on prête au monde de la tique un régime de signification qui n’est pas le sien, et notamment un système actantiel et prédicatif (prédateur, proie et « guet ») qui participe d’un autre monde que le sien, alors son monde sémiotique spécifique restera inaccessible et inconnaissable.
Le monde de la tique participe donc d’un régime de signification qui lui est propre, et le moindre faux pas dans la description peut compromettre l’interprétation, notamment par projection anthropomorphe. Certes, il faut se refuser à l’imaginer en train de « guetter sa proie », mais même également d’attendre une occasion : il est impossible de lui prêter une attente, et plus encore quelque patience ou impatience que ce soit, dès lors que son « instant » d’avant la chute peut durer presque indéfiniment, d’une seconde à plusieurs années, sans qu’on puisse repérer de modification de son comportement, autrement dit sans qu’elle en soit de quelque manière affectée.
Quant à l’occasion, il est vrai que le passage à proximité d’un mammifère est une conjoncture positive, mais seulement pour l’observateur humain, car la tique est bien loin de pouvoir calculer les chances de réalisation d’une conjoncture parmi les différents événements possibles ! Comme Uexküll le précise, la tique peut se laisser tomber sur un corps qui se révèle froid, et elle remonte alors se percher en hauteur ; le cycle recommence autant de fois que nécessaire, jusqu’à ce qu’elle tombe sur un corps chaud. Mais elle n’identifie jamais une « occasion », elle perçoit seulement la différence entre corps chauds et corps froids, et c’est le régime de signification de son Umwelt qui déclenche le processus de pompage du sang dans le cas des corps chauds, ou le processus de remontée en hauteur en cas de corps froids.
L’éthosémiotique animale commence donc par une prudente humilité et une exigeante ascèse interprétative, car la condition subjectale de construction des Umwelten implique un véritable zoomorphisme de l’analyse.
Le statut sémiotique des interactions peut être rapproché, en raison de leur rôle d’actualisation et de réactualisation des schèmes stabilisés et de l’Umvelt, de celui des énonciations. L’Umwelt d’un être vivant est d’abord constitué, avec une stabilité qui permet de le reconnaître à chaque occurrence d’événement, grâce à un certain nombre de sélections figuratives, de régimes temporels et spatiaux, de schèmes de comportement, et de configurations thématiques figées. Chaque interaction réactualise tout ou partie de ces propriétés spécifiques en les adaptant aux circonstances immédiates (ici et maintenant), en les modifiant ou en les reconfigurant, tout comme dans la praxis énonciative. La différence principale entre une manifestation sémiotique par interactions et une manifestation sémiotique par énonciation tient au fait que dans le second cas les actants en présence sont déjà connus et définis (ce sont les actants personnels de l’énonciation, des sujets et des objets), alors que dans le premier, ce n’est pas le cas : nous n’avons affaire qu’à des interactants, l’un d’entre eux étant choisi comme centre de référence pour l’Umwelt.
- Note de bas de page 33 :
-
Définies par René Thom dans plusieurs de ses travaux, notamment dans Esquisse d’une sémiophysique, Paris, InterEditions, 1988.
Il nous faut d’abord renoncer à poser comme préalable une structure actantielle différenciée, et, a fortiori, quelque schématisation narrative prédéterminée que ce soit. Si toute existence est définie comme coexistence (entre les existants), alors la signification de la coexistence ne peut être construite qu’à partir des interactions et des interactants, dont on ne connaît pas encore les rôles respectifs, mais dont on peut caractériser le régime sémiotique propre à leurs interactions. Ce régime pourra se caractériser par un mode d’existence, dont la typologie doit être établie (cf. infra, 3.3.4, les modes « réel », « imaginaire », « vital » et « ludique »), par une tonalité thématique, modale et affective (dont la typologie doit également être établie, cf. infra, id.). La description de ces régimes de signification devra également mobiliser les notions de prégnances et de saillances biologiques33.
- Note de bas de page 34 :
-
Principalement Présences de l’autre. Essais de socio-sémiotique II, Paris, PUF, 1997, et Les interactions risquées, op. cit.
Les seules conditions initiales requises pour que des interactions aient lieu sont l’altérité et l’énergie : des existants multiples et différents, et des flux dynamiques partagés. L’énergie n’est autre que la force qui met en relation les interactants. Assimilable à la phorie greimassienne, elle peut se manifester de multiples manières : par des stimuli perceptifs, des affects ou des rythmes partagés, des mouvements, etc. L’hypothèse d’une altérité constitutive est étayée sur la plupart des travaux interactionnistes, et, dans le domaine sémiotique, sur ceux d’Eric Landowski34. Par définition, des relations entre des semblables strictement identiques ne donnent lieu à aucune autre interaction que leur stricte répétition ; les interactants sont donc, a minima, des existants porteurs d’une force, et leurs interactions ne sont pertinentes (c’est-à-dire porteuses d’effets de sens) que s’ils sont différents les uns des autres, c’est-à-dire dotés de propriétés qui font de chacun d’eux un autre pour tous les autres.
Telle est donc la seconde condition pour accéder à la signification du monde vivant sans faire appel ni à la téléologie ni à une typologie des signes : une sémiotique des interactions est posée comme préalable à l’analyse narrative et à la déclinaison des régimes de signification des mondes vivants. Les deux conditions réunies (la lecture rétrojective et modalisante, et la sémiotique des interactions et de leurs régimes de signification) fournissent un point de départ plausible pour une biosémiotique qui vise la signification des mondes vivants, et non les signes et les buts de la vie.
3.2. Des interactions sensibles et réflexives
Comme nous l’avons déjà suggéré, la téléologie est presque systématiquement associée, dans le domaine du vivant, à d’autres types de schèmes argumentatifs. Les finalités, en effet, si elles ne sont pas incarnées dans un démiurge, une divinité ou une essence spéculative comme la nature, peuvent être plus subtilement exprimées grâce à une hiérarchie des espèces, où les humains occuperaient la position dominante, ce qui légitimerait les explications anthropomorphiques du vivant. Finalisme, position dominante des humains et anthropomorphisation du vivant forment alors un ensemble cohérent, rejeté par la plupart des communautés scientifiques, mais sous-jacent ou explicitement sollicité dans les raisonnements biosémiotiques. C’est pourquoi, par exemple, Richard Dawkins ironise à propos de la hiérarchie entre les espèces au regard de l’évolution :
- Note de bas de page 35 :
-
Richard Dawkins, Il était une fois nos ancêtres. Une histoire de l’évolution, Paris, Robert Laffont, 2007, p. 173.
Il n’y a pas plus de sens — et, également, pas moins — à diriger notre narration de l’histoire évolutive en direction d’Homo sapiens plutôt qu’en direction de n’importe quelle espèce — par exemple la pieuvre, ou la panthère ou le séquoia. Un martinet exceptionnellement avisé, fier, de façon très compréhensible, d’avoir la capacité de voler, regardera le vol comme l’évident accomplissement ultime de la vie, et considèrera son espèce — ces spectaculaires machines volantes avec leurs ailes ramenées en arrière, qui restent parfois en l’air une année et même copulent en vol libre — comme l’acmé du progrès évolutif. Pour reprendre une fable imaginée par Steven Pinker, si les éléphants avaient la capacité d’écrire l’histoire, ils pourraient décrire les tapirs, les musaraignes éléphants, les éléphants de mer et les nasiques comme des premières tentatives pour emprunter l’axe principal de la route évolutive, franchissant les premières étapes mais, pour certaines raisons, n’achevant jamais complètement leur évolution : si près, mais encore si loin. Des éléphants astronomes pourraient se demander si, sur d’autres planètes, il existe des formes de vie extraterrestre ayant franchi le rubicond nasal et fait le bond vers la complète trompitude.35
- Note de bas de page 36 :
-
Relisons à cet égard, et sans autre commentaire, ce passage bien connu de Bernardin de Saint-Pierre, auquel quelques biosémioticiens se réfèrent parfois, sans sourire : « Il n’y a pas moins de convenance dans les formes et les grosseurs des fruits. Il y en a beaucoup qui sont taillés pour la bouche de l’homme, comme les cerises et les prunes ; d’autres pour sa main, comme les poires et les pommes ; d’autres beaucoup plus gros comme les melons, sont divisés par côtes et semblent destinés à être mangés en famille : il y en a même aux Indes, comme le jacq, et chez nous, la citrouille qu’on pourrait partager avec ses voisins. La nature paraît avoir suivi les mêmes proportions dans les diverses grosseurs des fruits destinés à nourrir l’homme, que dans la grandeur des feuilles qui devaient lui donner de l’ombre dans les pays chauds ; car elle y en a taillé pour abriter une seule personne, une famille entière, et tous les habitants du même hameau ». Bernardin de Saint-Pierre, Études de la nature, 1784 (ch. XI, section « Harmonies végétales des plantes avec l’homme »).
L’anthropomorphisme fait souvent sourire, mais il a pourtant dirigé les réflexions sur le monde vivant pendant des siècles, et suscité systématiquement des raisonnements téléologiques36.
- Note de bas de page 37 :
-
Il en résulte concrètement que les biologistes consultés considèrent que la plupart des problèmes et modèles actuellement discutés par les biosémioticiens ont été traités, résolus ou abandonnés dix ou vingt ans plus tôt par les biologistes eux-mêmes.
Pourtant, le rejet massif et sans discussion de la téléologie, au seul nom des principes, n’est pas la solution la plus productive. Au contraire, ce sont les critiques et les remaniements, le détail des controverses et des consensus qui permettent, tout au long de l’histoire de cette conception du monde vivant, de dégager progressivement des lignes théoriques et des principes de méthode susceptibles de nourrir un dialogue utile et productif entre la biologie et la sémiotique. A cet égard, la sémiotique ne peut pas, comme c’est souvent le cas encore actuellement, se cantonner à l’exploitation des failles, des manques, des ignorances et des incertitudes dans les recherches et les théories biologiques : cela ne déboucherait pas sur un dialogue très diplomatique, et surtout ne produirait qu’une sémiotique éphémère, vite obsolète, au fur et à mesure des progrès accomplis par les recherches biologiques37.
- Note de bas de page 38 :
-
Cette remarque est très caractéristique des aléas du dialogue entre la sémiotique et les sciences du vivant, et de la vigilance qu’il impose. En effet, cette tentation anthropomorphe sera de moins en moins acceptable ; elle l’était encore il y a quelques années mais les recherches et les résultats en matière de sensibilité animale se développent aujourd’hui massivement et il ne sera bientôt plus possible, sur ce sujet, de faire de l’anthropomorphisme une tentation acceptable ou compréhensible, car le fonctionnement de la sensibilité animale sera connu, disponible, et difficile à ignorer !
- Note de bas de page 39 :
-
Jacques Dewitte, « La vie est sans pourquoi. Redécouverte de la question téléologique », Revue du MAUSS, 2008, 31, 1, pp. 435-465.
- Note de bas de page 40 :
-
Robert Spaemann, « Téléologie de la nature et action humaine » (1978), traduit de l’allemand par Pierre Destrée et Jacques Dewitte, Phénoménologie et philosophie de la nature. Études phénoménologiques, 23-24, Bruxelles, Ousia, 1996. Thure von Uexküll, « Critique des présupposés contemporains en matière de philosophie de la nature et de notre attitude à l’égard du problème de la téléologie » (chapitre IV de Der Mensch und die Natur, 1953), traduit de l’allemand par Georges Thinès, ibid. Hans Jonas, « Les fondements biologiques de l’individualité » (1968), traduit de l’anglais par Danielle Lories, ibid. Adolf Portmann, « L’autoprésentation, motif de l’élaboration des formes vivantes » (1958), traduit de l’allemand par Jacques Dewitte, ibid.
L’une des difficultés que nous rencontrons à cet égard est celle d’un traitement non anthropomorphe de la sensibilité au sein des mondes vivants38. Dans un article de référence, « La vie est sans pourquoi », Jacques Dewitte a examiné il y a une dizaine années les discussions conduites par les biosémioticiens et philosophes du vivant39. Il évoque notamment les travaux de Robert Spaemann, Thure von Uexküll (le fils de Jakob von Uexküll), Hans Jonas et Adolf Portmann40. Il en résulte quelques conséquences qui nous permettent de préciser le type de biosémiotique que nous cherchons à circonscrire et définir.
3.2.1. Sensibilité et figurativité
- Note de bas de page 41 :
-
En écho de la corrélation entre les images-perceptions et les images-actions de von Uexküll.
Un premier consensus semble se dessiner, qui conforte notre référence à la sémiotique des interactions : un monde vivant, à la différence de tout autre domaine de l’existence, est peuplé d’existants sensibles, et c’est donc un monde vécu (Uexküll, père et fils). Un monde vécu est nécessairement (par voie de conséquence) un monde organisé autour d’un centre de référence, qui est plus précisément dans ce cas un centre de sensibilité et d’activité41, pour lequel les interactions sont des actions qui affectent la sensibilité, et des états sensibles qui affectent l’action.
Cette relation réciproque, activée dans le processus des interactions, a notamment pour conséquence que si, sur l’une des deux faces, sensibilité ou action, les interactions modifient les propriétés de l’être vivant, il se ressent néanmoins, au titre de l’autre face, identique à lui-même : l’action le transforme, mais sa sensibilité le confirme dans son identité ; ses états sensibles sont modifiés, mais dans l’action il s’affirme identique à lui-même. Si les deux faces sont transformées simultanément, il y a alors un risque de désorientation et de perte d’identité. Nous reviendrons sur cette question. Cette relation réciproque, entre sensibilité et activité, est la signature du vivant, car elle instaure une boucle rétroactive qui induit et stabilise chez l’être vivant l’éprouvé d’une identité (en termes anthropomorphes, on dirait « le sentiment d’identité »).
Indépendamment de toute interaction, le centre de référence de l’Umwelt se définit a minima comme sensibilité et activité. Dès qu’il s’actualise dans une ou des interactions, la boucle de rétroaction est mise en œuvre, et il devient alors un centre de réflexivité à partir duquel se construit l’identité à soi-même de l’organisme vivant : en ce centre, ce dernier est stimulé par son milieu, configure ces stimuli en signaux ou en d’autres expressions sémiotiques, comme des activités pratiques, qui elles-mêmes influent sur le milieu, et le cycle recommence et renforce l’identité de l’existant central.
- Note de bas de page 42 :
-
Adolf Portmann, « L’autoprésentation, motif de l’élaboration des formes vivantes », art. cit.
On peut alors ajouter à ce centre sensible, actif et réflexif, une capacité d’« auto-présentation » figurative, comme le fait Portman42. Il soutient par exemple que l’identité des organismes vivants se manifeste également par leurs formes sensibles — notamment des propriétés plastiques, des formes, des couleurs, des bruits, des cris spécifiques, etc. Il interprète ces formes sensibles de l’auto-présentation comme des expressions d’une finalité propre au monde de la vie, mais l’appel aux causes finales n’ajoute rien de significatif à cette propriété.
En effet, les formes sensibles de chacun des êtres vivants (par exemple les couleurs et graphismes sur les ailes du papillon) sont des manifestations figuratives, adressées à la fois à soi-même, aux congénères et aux autres espèces. Elles ont donc une double destination sensible : vers la propre sensibilité de l’entité vivante centrale, et vers celle de ses » autres ». Il n’est pas utile ici de distinguer l’adresse aux congénères et l’adresse aux autres espèces, car Uexküll a montré qu’au sein d’un Umwelt, la frontière entre ces deux types d’« autres » est mobile et poreuse. L’identité figurative de l’espèce, ou même de l’individu, composée d’un plan de l’expression — des formants et d’un plan du l’identité elle-même —, est ainsi donnée à percevoir et à ressentir, à soi-même, pour confirmation de l’éprouvé d’identité, et aux autres, pour infléchir leur participation à l’interaction. A ce titre, elle participe aux boucles réflexives évoquées plus haut.
En outre, ce sont à la fois des manifestations sensibles adressées, et des énonciations de l’appartenance à l’espèce. Ces manifestations figuratives participent aux interactions conçues comme des réactualisations énonciatives des propriétés d’un monde vivant donné. Si on part du principe que ce sont les interactions qui sélectionnent les propriétés pertinentes (c’est-à-dire participant à un même régime de signification) aussi bien de l’organisme central que de tous les autres participants, alors l’enjeu d’une affirmation figurative de l’identité est l’équivalent d’une prise de pouvoir au sein de l’Umwelt : l’un des interactants affirme qu’il est maître du jeu, et que la signification des interactions ne pourra se construire que de son point de vue. Mais chaque être vivant en interaction est soi-même un centre de sensibilité-activité-réflexivité pour tous les autres, et réciproquement. Il s’ensuit que les expressions figuratives de l’identité, dans l’apparaître des êtres vivants, sont la contrepartie de leur propre sensibilité subjectale : ils anticipent ainsi la sensibilité des autres participants et lui donnent une clé d’interprétation figurative de ce qui se passe ou va se passer, dans les pratiques en cours dans l’Umwelt.
L’identité figurative apparaît alors comme un foyer de tactiques véridictoires potentielles : affirmer ce qu’on est, annoncer les phases pratiques de l’interaction, ou bien les dissimuler, les remplacer par d’autres qui sont des annonces erronées. En offrant ainsi à l’autre matière (figurative) à sentir, à réagir, et à mettre en œuvre ou à adapter des schèmes de comportement, l’être vivant ne se contente pas d’interagir avec l’autre : il l’incite, il le provoque, il le dissuade, il lui fait baisser la garde, il le prend par surprise, en somme, il l’influence ou le manipule. La co-sélection des expressions figuratives de chaque espèce et de chaque individu est de ce fait à la fois le fruit des interactions, une condition de leur fonctionnement régulé, et un instrument d’influence et de manipulation. Comme tous les « autres » ont également une identité figurative, l’auto-présentation ouvre également la possibilité, à tout moment, à des contre-tactiques et des basculements de point de vue entre les interactants.
Nous pourrions, par facilité, parler ici de « communication » entre êtres vivants, mais la généralisation du concept et des modèles de la communication dans le monde vivant serait un de ces actes théoriques inopportuns que, tout comme le raisonnement téléologique, nous nous efforçons ici de critiquer et de contenir. Nous sommes ici confrontés, à l’évidence, à une interaction entre au moins deux entités vivantes qui se ressentent mutuellement comme sensibles et susceptibles de reconnaître chez l’autre des formes sensibles iconisées. L’apparence figurative est l’icône de l’espèce, peut-être même, dans bien des cas, l’icône de l’état actuel d’activité et de réceptivité sensible de l’individu. L’apparaître en est l’énonciation. Comme nous l’avons suggéré, l’auto-présentation figurative est en effet l’un des actes constitutifs des énonciations pratiques, c’est-à-dire un acte adressé à la fois à l’autre et à soi-même. Il en résulte que parmi les interactions qui caractérisent les mondes vivants, les plus spécifiques relèvent de la manipulation, de l’influence ou de l’incitation, en même temps que de l’affirmation de soi.
3.2.2. Sensibilité et autoconservation
Un autre point de consensus apparaît : dès lors qu’on ne la considère plus comme une finalité, l’autoconservation du vivant résulte d’une résistance à tout ce qui pourrait compromettre la poursuite du cours de l’existence. Mais il faut alors bien circonscrire ce qui, spécifiquement pour les êtres vivants, se conserve. Il ne suffit pas en effet d’invoquer la persistance de l’existence, ni même la résistance à ce qui serait susceptible de la compromettre : la montagne persiste, malgré l’érosion, et cela n’en fait pas pour autant un être vivant ; éventuellement un animé, pour l’animisme et le totémisme, mais pas un vivant. On pourrait invoquer les échelles de temps, et avancer que la montagne finit par se réduire à un plateau, mais cette observation triviale ne résisterait pas à la confrontation avec l’échelle de temps de la disparition et du remplacement des espèces vivantes.
- Note de bas de page 43 :
-
Hans Jonas, « The burden and blessing of mortality », Philosophische Untersuchungen und metaphysische Vermutungen, Francfort, Insel Verlag, 1992, p. 87.
Hans Jonas fait remarquer en revanche que l’autoconservation s’oppose, non pas seulement à des interventions délétères extérieures, mais à l’un des principes mêmes de la vie : le métabolisme propre aux êtres vivants est un type spécifique d’interaction, reposant sur des échanges de matières et d’énergie avec leurs milieux, qui paradoxalement suscite et conforte leur identité : elle perdure en effet malgré le renouvellement cyclique complet de leurs propres constituants. Si on met entre parenthèses quelques extrapolations spéculatives, son analyse consiste à projeter sur le métabolisme une structure narrative élémentaire, celle de l’actant et de l’antactant, une structure polémique dont il résulte que la vie en tant que processus comporte en interne à la fois une direction de contre-persistance (le renouvellement complet de l’organisme) et une direction de persistance (le maintien de l’identité à soi-même)43.
Il complète alors le mécanisme de renouvellement métabolique par une affirmation de la vie contre la mort. Une biosémiotique plus prudente peut se satisfaire du principe de la persistance, et de la résistance aux assauts et aux aléas de la contre-persistance. Il n’est pas même nécessaire de faire appel à des rôles actantiels définis, puisque les tensions modales, entre pouvoir et ne pas pouvoir notamment, suffisent à rendre compte de cette résistance. La distribution des équilibres et déséquilibres, des confrontations et des ajustements, tout au long du processus existentiel, suscitera alors, voire stabilisera des schèmes syntagmatiques récurrents et identifiables. Les formes syntagmatiques de la persistance offrent dans cette perspective une diversité suffisante pour que la construction de leur signification dégage des systèmes de valeurs et des régimes de sens, sans qu’il soit nécessaire d’attribuer aux êtres vivants un « intérêt » et un « attachement » à la vie même.
Mais la solution est encore incomplète car cette identité maintenue malgré le renouvellement métabolique n’apparaît qu’aux yeux d’un observateur extérieur, et elle pourrait tout aussi bien être constatée pour des systèmes physiques en mutation, des objets qui s’usent ou des montagnes qui s’érodent. Le raisonnement qui précède complique la manière de poser le problème, mais sans le résoudre ! Il manque ici la dimension subjectale que nous avons déjà reconnue comme caractéristique des mondes vivants. Il manque aussi à ce raisonnement le fait que l’être vivant ne se résume pas à son métabolisme, et que ses interactions avec le milieu impliquent nécessairement une sensibilité. En résumé, l’autoconservation doit également assurer non seulement la persistance de la sensibilité, mais aussi et surtout celle de l’éprouvé sensible de l’identité.
Hans Jonas a bien vu la difficulté : il commence par admettre lui aussi que la sensibilité fait partie de l’existence des êtres vivants (cf. supra, en tant que centres réflexifs de sensibilité-activité), et il propose un simple déplacement du principe d’autoconservation : l’être vivant sentant se maintient en vie comme être sentant, et pas seulement comme être métabolisant. Dans le conflit entre les deux directions de la persistance et de la contre-persistance, ce serait donc le centre de réflexivité qui serait l’arbitre, et qui animerait la résistance de la première contre la seconde. Dès lors, l’autoconservation serait principalement non pas celle de l’existence objective mais celle d’une existence vécue, sensible et subjectale.
- Note de bas de page 44 :
-
Jean-Paul Petitimbert, « Lecture d’une pratique et d’une interaction : l’hésychasme orthodoxe », Actes Sémiotiques, 118, 2015 ; id., « Amor vacui. Le design d’objets selon MUJI », Actes Sémiotiques, 121, 2018. Paolo Demuru, Essere in gioco, Bologne, Bononia University Press, 2014 ; id., « Malandragem vs Arte di arrangiarsi : Stili di vita e forme dell’aggiustamento tra Brasile e Italia », Actes Sémiotiques, 118, 2015. Eric Landowski, « L’épreuve de l’autre », Sign Systems Studies, 34, 2, 2008. Maria Cristina Addis, « Forme d’aggiustamento. Note semiotiche sulla pratica dell’aikido », in Ana Cl. de Oliveira (éd.), As Interações sensíveis. Ensaios de sociossemiótica, São Paulo, Estação das Letras e Cores, 2013. Pierluigi Cervelli, « Fallimenti della programmazione e dinamiche dell’aggiustamento. Sull’autoproduzione dello spazio pubblico », in As Interações…, op. cit. Diana Luz Pessoa de Barros, « Les régimes de sens et d’interaction dans la conversation », Actes Sémiotiques, 120, 2017.
Ce déplacement auquel nous souscrivons sans hésiter est décisif car il nous rapproche plus encore de la sémiotique des interactions telle qu’elle se développe aujourd’hui dans le champ socio-sémiotique : comme le rappellent avec insistance Landowski et bien d’autres44, les événements de la sensibilité ne sont pas seulement des supports et des occasions pour les interactions, mais des enjeux pour les régimes de sens. Les interactions sensibles ont en effet comme principaux résultats des métamorphoses et des reconfigurations de la sensibilité elle-même, ce qui relance le problème posé : sur quoi s’appuyer si l’organisme vivant est en transformation permanente à la fois de son métabolisme et de sa sensibilité ? Comme indiqué plus haut, il faut alors s’appuyer sur les boucles de rétroaction entre action (à ce niveau d’analyse, il s’agit des actions du métabolisme) et sensibilité.
Par conséquent, ce qui se conserve de plus précieux dans cette autoconservation, c’est bien l’éprouvé d’identité qui résulte de ces boucles de rétroaction entre activité et sensibilité : les individus continuent à s’éprouver comme identiques à eux-mêmes, et comme appartenant à la même espèce que leurs congénères. Dans les mondes vivants, la principale reconfiguration est celle qui, en remaniant les propriétés figuratives et sensibles co-sélectionnées, assure la réactualisation permanente non seulement de la coexistence des interactants et de leurs identités respectives, mais aussi et surtout la pérennité de l’éprouvé de l’identité à soi-même.
Cet éprouvé identitaire est en effet au cœur de la différenciation des régimes sémiotiques : c’est à partir de lui que les êtres vivants peuvent également éprouver le rapport entre soi et autrui, éprouver les risques que ce rapport fait encourir au soi, y projeter des schèmes pratiques, des scènes thématiques, faire varier ou interchanger les rôles, etc. Nous verrons tout à l’heure que la théorie des tonalités chez von Uexküll aboutit à un résultat comparable : les tonalités sont en effet pour l’éthologue l’équivalent de nos régimes sémiotiques, organisés autour d’une scène pratique qui participe le plus souvent des prégnances biologiques ; mais surtout, chacune caractérise à sa façon la manière dont l’animal s’éprouve lui-même à un moment donné, et dont il sélectionne les « autres » pour jouer un rôle dans sa propre scène thématique.
- Note de bas de page 45 :
-
Milieu animal et milieu humain, op. cit., pp. 134-135.
- Note de bas de page 46 :
-
Georges Brassens, dans la chanson Le Gorille, évoque des prolongements plus scabreux.
Ce fonctionnement est d’autant plus convainquant que le choix du partenaire est incongru : Uexküll raconte par exemple qu’un jeune butor mâle avait choisi son directeur de zoo comme partenaire sexuel45. Le partenaire en question se montrant peu coopératif et s’étant par précaution durablement tenu à l’écart, le butor a alors accepté une femelle de son espèce, qui a donné le jour à leur progéniture. Mais, croyant que cet heureux dérivatif avait fait disparaître les fantaisies trans-espèces du mâle, le directeur du zoo s’est à nouveau montré à lui sans précaution : le butor mâle a aussitôt chassé la femelle et fermement invité le directeur à prendre sa place dans le nid. Uexküll ne dit pas comment l’affaire s’est conclue46.
Une telle situation où un animal choisit un interactant incongru, pour toutes sortes de pratiques, n’est pas une rareté, et elle est très probante : on voit bien ici que la tonalité sexuelle ne se construit pas à partir de l’identité (d’espèce, de sexe, d’apparence figurative) de l’autre, mais à partir de la manière dont il s’éprouve lui-même (l’éprouvé d’identité du soi), et de la construction tonale et thématique qu’il projette sur l’autre, quel qu’il soit, en se fondant sur cet éprouvé décisif. Le « bon sens » (anthropomorphe ?) voudrait que le butor procède d’abord à une identification de l’autre comme partenaire sexuel possible. Mais la prégnance biologique exerce sa pression non pas d’abord sur la perception qu’il a de l’autre, mais directement sur la manière dont il s’éprouve en tant que soi-même.
3.2.3. Sensibilité et perspectivisme
- Note de bas de page 47 :
-
Maurice Merleau-Ponty, « Le concept de nature », Résumés de cours, Paris, Gallimard, 1968, p. 136.
L’anthropomorphisme, tout comme la téléologie, n’est pas condamné par principe, mais doit être soumis à un examen critique afin de susciter des positions alternatives. L’exclure systématiquement nous ferait courir le risque de reconstituer un point de vue anthropomorphe dominant et surplombant, celui de l’analyste, qui serait alors inaccessible à la critique théorique et méthodologique. Si l’anthropomorphisme ne peut pas être oublié ou neutralisé, il est néanmoins possible d’en contrôler et réduire les effets en s’inspirant notamment des réflexions de Merleau-Ponty : « Toute zoologie suppose de notre part une Einfühlung méthodique du comportement animal, avec participation de l’animal à notre vie perceptive, et participation de notre vie perceptive à l’animalité »47.
L’anthropomorphisme critique, à la manière de l’anthropologie « cannibale » de Viveiros de Castro, implique donc la réciproque : une animalisation de l’observateur humain, devenu capable d’aller et venir, de manière critique, entre le point de vue humain et le point de vue animal, en entretenant une certaine perméabilité entre les deux types de subjectalités. L’empathie (Einfühlung) n’explique pas grand-chose, mais indique suffisamment la voie méthodologique : se mettre à la place de l’autre, sans perdre notre identité, pour qu’il puisse se mettre en retour à notre place, sans perdre la sienne.
- Note de bas de page 48 :
-
Dominique Lestel, « De Jacob von Uexküll à la biosémiotique », in Jacob von Uexküll, op. cit., pp. 17-21.
Nous avons déjà indiqué que les mondes vivants sont d’abord organisés autour d’un organisme central (le centre sensible, actif et réflexif de l’Umwelt), et que, de son côté, l’observateur devait adopter cette position comme centre d’orientation épistémique. Cette conception est aujourd’hui radicalisée par les éthologues, et notamment par Dominique Lestel, dans son introduction à la traduction française de Milieu animal et milieu humain de Jacob von Uexküll, où il propose comme modèle de méthode éthosémiotique la démarche des artistes : « La grande différence entre les artistes et l’écrasante majorité des spécialistes universitaires de l’animal réside en effet dans la capacité des premiers à s’animaliser pour pénétrer dans le monde étrange et merveilleux des Umwelten non humains »48.
- Note de bas de page 49 :
-
Loc. cit., p. 21.
- Note de bas de page 50 :
-
Il va de soi que dans bien des cas, notamment en raison des différences de taille (par exemple avec la fourmi) et des échelles d’analyse (comme pour la bactérie), ce perspectivisme ne dépassera pas le stade d’une expérience de pensée.
L’animalisation, que préconisait déjà Merleau-Ponty, pour neutraliser les effets du point de vue anthropomorphe de l’observateur, ne doit pas être de pure forme, mais bien réelle, car elle doit s’affranchir, précise Lestel, des barrières hygiéniques entre les espèces. Il évoque même une intoxication interspécifique49, qui serait en quelque sorte le point ultime de l’anthropomorphisme critique, devenu zoomorphisme. Pour prolonger également jusqu’à son terme le parallèle avec l’ethnologie et l’anthropologie, il s’agit de se faire autre tout en restant soi-même, en particulier pour pouvoir reconstituer en position subjectale ce que sont les « autres de l’autre », c’est-à-dire ici, les autres de l’animal, dont l’observateur doit s’efforcer de faire partie, pour accéder à la signification du monde animal50 : parallèlement à l’anthropologie perspectiviste défendue par Viveiros de Castro, l’anthropomorphisme critique déboucherait alors sur une éthosémiotique perspectiviste.
3.2.4. Récapitulons
Prendre en compte les formes et rôles de la sensibilité dans les mondes vivants implique par conséquent :
- Note de bas de page 51 :
-
Si la signification des formes d’existence (les régimes de signification qui leur sont associés) ne réside pas dans leur finalité, alors elle se loge dans la diversité des schèmes syntagmatiques de leur persistance. C’est précisément aussi le principe directeur d’une sémiotique des formes de vie, telle qu’elle est présentée dans J. Fontanille, Formes de vie, Liège, Presses Universitaires de Liège, 2015.
-
de considérer ces mondes comme vécus par des êtres sensibles, l’un d’entre eux étant choisi, dans chaque monde vivant, comme centre de sensibilité-activité ;
-
de prendre en compte les boucles de rétroaction entre sensibilité et activité, d’où émergent la réflexivité, et notamment l’éprouvé de l’identité à soi-même ;
-
de concevoir l’autoconservation du vivant comme une propriété intrinsèque de la vie saisie dans son cours, y compris la conservation de la sensibilité, et la variation des régimes de signification de ce cours, comme un effet des différentes formes sensibles que peuvent prendre les schèmes syntagmatiques de la persistance51 ;
-
de comprendre la persistance de la vie comme une résistance à la contre-persistance, dont le noyau opérateur est l’éprouvé de l’identité à soi-même ;
-
d’accorder à la manifestation figurative et sensible des êtres vivants une participation aux actes d’énonciation et d’actualisation des mondes vivants ;
-
de développer une éthosémiotique perspectiviste, en s’appuyant à la fois sur un anthropomorphisme critique et une animalisation méthodologique.
3.3. La sémiotique tonale de Jacob von Uexküll
Nous avons déjà mentionné l’éthosémiotique de Jacob von Uexküll, notamment pour sa conception de l’Umwelt en tant que domaine d’interactions organisé autour de ce qu’il appelle un « sujet », et que nous avons redéfini comme un centre réflexif de sensibilité et d’activité. Autre évocation de son œuvre, plus développée que les précédentes, la présente section a trait principalement à la question de la schématisation des comportements pratiques et de leurs usages comme tonalités des Umwelten.
- Note de bas de page 52 :
-
J. von Uexküll, op. cit., pp. 130-131.
Globalement, la schématisation résulte d’abord des co-sélections de propriétés perceptives et kinésiques qui caractérisent chaque Umwelt. Par exemple, un choucas reconnaît dans son Umwelt un autre choucas uniquement à partir de sa forme mouvante et de sa couleur noire : il aura les mêmes comportements si on lui présente un chiffon noir en mouvement52. Ensuite, si le même choucas est mis en présence d’un chat qui tient un autre choucas dans sa gueule, puis du même chat tenant un chiffon noir, dans les deux cas le premier choucas se met en position d’attaque. Des traits plastiques sont sélectionnés, et ils suffisent à déclencher la posture sensori-motrice qui prépare à la pratique induite par la situation perçue. On peut dire en ce cas que le choucas entre en interaction en mettant en œuvre un schème de comportement.
Les schèmes en question sont à chaque nouvelle occurrence réactualisés, en fonction des individus, des circonstances spatiales, temporelles et actorielles, et plus généralement de la situation où se trouve l’Umwelt au moment des événements en cours. Nous avons déjà proposé de considérer ces réactualisations comme des formes d’énonciation pratique, mais il reste à examiner de plus près comment on passe des schèmes potentiels, considérés comme caractéristiques des espèces, à leurs occurrences particulières, telle qu’elles sont réalisées par des individus.
3.3.1. La schématisation pratique
A propos de la tique qui reconnaît le passage d’un mammifère et qui se laisse tomber sur lui pour aspirer du sang, Uexküll évoque quatre perceptions successives (l’odeur de l’acide butyrique émis par les follicules sébacés des mammifères, la chaleur du corps, le contact tactile du pelage ou de la fourrure, l’identification du sang en tant que liquide chaud), et quatre actions (se laisser tomber, chercher une zone de faible densité pileuse, perforer et pomper le sang).
- Note de bas de page 53 :
-
Ibid., pp. 40-41. Uexküll compte seulement trois arcs réflexes successifs, mais dans sa description, reconstituée à partir de plusieurs développements disjoints dans son livre, on compte bien quatre actions intercalées avec quatre perceptions.
Uexküll considère que la succession entre une perception et une action forme un arc réflexe isolable, et la séquence qu’il décrit n’est de ce fait qu’une suite d’arcs réflexes53. Pourtant, l’enchaînement syntagmatique qui doit assurer le succès de la série tout entière obéit nécessairement à une autre règle que celle de l’arc réflexe, puisque, comme le précise l’auteur, dans l’arc réflexe l’action « éteint le signe perceptif » (p. 39). En outre, la liaison entre les quatre phases successives reste contingente : le parcours à accomplir peut être interrompu à chaque phase, si les conditions perceptives et actionnelles ne sont pas remplies. Il ne s’agit donc pas d’une loi transcendante imposée de l’extérieur (comme une finalité, par exemple), mais tout simplement d’une régulation interne d’interactions en série.
Uexküll invoque à cet égard d’abord une « construction intégrative » (p. 39), puis un « plan » : « Il appert que les trois cercles fonctionnnels se succèdent conformément à un plan » (p. 40). Il précise plus loin que ces cercles se suivent en « se relayant les uns les autres » (p. 41). Intégration, plan ou relais, le nom donné au problème ne suffit pas à le traiter. Ce n’est que dans la suite de l’ouvrage qu’il évoquera des « schèmes », dans le sens kantien du terme, c’est-à-dire comme position intermédiaire entre la conceptualisation et l’intuition sensible. Chaque phase est une interaction, et l’ensemble de ces interactions successives est lui-même une interaction globale entre deux interactants, organisée par un schème syntagmatique d’interaction. En d’autres termes, on pourrait dire que la forme syntagmatique globale de l’interaction est soumise à la pression d’une prégnance biologique, alors que chaque phase d’interaction correspondrait à une saillance. La seule régulation globale qui s’impose à la tique, c’est la persistance de l’espèce, dont elle n’a aucune connaissance ou conscience, et il faut donc prévoir un niveau d’intégration des perceptions et des actions qui soit plus proche de la sensibilité animale : faute de mieux, nous choisissons celui du schème, et c’est le schème de la préparation de la ponte qui est ici sélectionné : si, et seulement si elle accomplit la séquence toute entière elle pourra pondre les œufs fécondés qu’elle porte déjà.
- Note de bas de page 54 :
-
La distinction proposée par le Groupe μ dans les Principia semiotica (op. cit.), entre anasémiose (fondée sur une perception) et catasémiose (fondée sur une action) va dans le même sens car elle permet d’articuler les deux types de sémioses comme des modes d’existence successifs et différents de la signification : la sémiose anasémiotique actualise la sémiose catasémiotique associée, qui potentialise une perception de l’action, et donc une signification anasémiotique, etc.
Du point de vue de l’observateur humain, les perceptions et les actions sont articulées en séquences canoniques porteuses du sens global de la pratique animale, chacune étant précisément située en une phase déterminée. Mais, de son point de vue, la tique n’a affaire qu’à un schème d’interaction, c’est-à-dire à une pression somatique et sensible qui l’incite à enchaîner les interactions entre perceptions et actions. Chaque perception actualise une action, qui elle-même potentialise la perception suivante, et ainsi de suite54. Uexküll invoque des « signes perceptifs » et des « signes actionnels », mais ils ne reçoivent leur statut sémiotique (comme pour des morphèmes ou des syntagmes dans une phrase), et surtout leur contenu pertinent, que de l’ensemble de la séquence. Ce sont donc, stricto sensu, non pas des signes, mais des figures, qui ne prennent leur valeur que dans le réseau de toutes les autres figures distribuées dans la séquence, et en raison de leurs positions syntagmatiques respectives : la moindre neutralisation ou permutation annule ou modifie cette valeur. Une fois que la séquence régulatrice est reconnue par l’analyste, elle acquiert une stabilité suffisante pour se prêter à toutes sortes de manipulations : saisie à partir de la fin, elle se donne comme un « destin » ; saisie à partir de son début, elle peut passer pour un « projet » ; projetée toute entière sur d’autres individus et d’autres groupes de la même espèce, elle peut donner lieu à des prévisions, sinon à des prédictions.
Encore une fois, l’éthosémiotique de von Uexküll conduit à une sémiotique de la signification des interactions et de leurs processus, et la sémiotique des signes et de leur typologie à laquelle il fait appel pour commencer n’est qu’un habillage terminologique des acquis de la biologie et de l’éthologie animale, un préambule pour une autre construction sémiotique.
- Note de bas de page 55 :
-
Pour la tique, par exemple, il n’y a pas de « mammifère hôte », mais un paquet de propriétés sensibles nécessaires et solidaires : odeur, chaleur, texture.
Les pratiques animales seraient donc régulées par des schèmes syntagmatiques, dont le contenu sémantique global serait dans la plupart des cas celui de prégnances biologiques. La schématisation présuppose elle-même deux opérations nécessaires : l’orientation subjectale qui fixe le centre de référence pour toutes les interactions jugées pertinentes dans l’Umwelt, et la co-sélection des propriétés pertinentes de l’organisme vivant et de son milieu55, qui ont pour contenu les saillances biologiques propres à une espèce. En bref, comme un Umwelt est caractéristique d’une espèce ou d’un groupe d’organismes, l’orientation subjectale, la sélection et la schématisation sont trois opérations complémentaires qui participent à la caractérisation de l’Umwelt de chaque espèce animale.
3.3.2. Les tonalités
Uexküll distingue successivement deux modes de fonctionnement de ce qu’il appelle des « signes » : en tant que signes proprement dits, et en tant que « tonalités ». Mais dès qu’il aborde les tonalités, la notion de signe disparaît quasiment de son argumentaire et elle est remplacée par celle d’« image » (image perceptive, image actionnelle), ce qui équivaut à nos figures (cf. supra). Les tonalités ne sont donc pas des modes de fonctionnement des signes, mais au contraire des régimes sémiotiques de niveau supérieur aux signes, qui s’imposent dès que les signes ne sont plus traités que comme des figures. Les figures sont soit des expressions qui recevront une part du contenu des tonalités, soit des contenus qui trouveront leur expression dans l’organisation observable des tonalités.
La tonalité elle-même n’est ni un signe ni une figure mais une configuration complexe qui caractérise directement les interactions, et pas l’organisme ou son espèce. Elles sont donc d’ordre écologique. Une tonalité se définit d’abord en tant que dominante thématique d’une pratique : on prend, on échange, on se protège, on se reproduit, on se déplace, etc. Mais la tonalité peut également correspondre à un affect, par exemple la peur ou la répulsion. Cette dominante thématique ou passionnelle implique une scène comprenant des rôles et un prédicat principal, qui en général donne son nom à la tonalité, par exemple une « tonalité d’alimentation » (soit : prédicat ‘alimenter’, actant 1 ‘aliment’ et actant 2 ‘alimenté’).
Toutefois, ce n’est pas la situation de l’Umwelt qui dicte la tonalité, mais le contraire : c’est la tonalité qui donne sens à l’Umwelt. Et pour preuve : un même Umwelt, pour un même animal, en interaction avec les mêmes éléments du milieu, peut être actualisé sous plusieurs tonalités alternatives, voire exclusives l’une de l’autre : le bernard-l’hermite, face la forme creuse plus ou moins cylindrique d’une anémone de mer, peut interagir avec elle en la traitant soit comme habitat s’il n’a pas de coquille, soit comme protection contre les prédateurs s’il en appréhende l’approche, soit comme nourriture, s’il est affamé. Les figures perceptives ne changent pas, seules les thématiques pratiques alternent : se loger, se protéger ou se nourrir. Il en résulte trois scènes et trois systèmes de rôles différents, qui sont, au moins pour l’enveloppe de l’anémone, incompatibles : elle ne peut pas à la fois être dévorée et servir de protection ou de logement. La tonalité sélectionnée détermine à la fois l’Umwelt, le rôle de l’anémone et celui du bernard-l’hermite, mais elle n’est elle-même sélectionnée que par l’éprouvé actuel du bernard-l’hermite : en tant qu’habitant errant, en tant que proie potentielle, en tant qu’affamé. L’éprouvé en question n’est plus seulement celui d’une identité générale, mais celui d’une configuration contingente, liée aux circonstances hic et nunc, qui n’est qu’une facette provisoire de l’identité générale.
Trois schèmes syntagmatiques sont à l’œuvre, correspondant à trois prégnances biologiques différentes. Chacun est actualisé par l’organisme vivant en fonction des caractéristiques de sa situation présente, et surtout de la pression d’une prégnance qui s’exerce hic et nunc, et dont la manifestation sémiotique est une tonalité. Ce type d’alternatives foisonnent, pour la plupart des organismes vivants, et sont même susceptibles d’engendrer d’apparentes « erreurs », comme nous l’avons déjà signalé : un choucas peut par exemple, par ses mouvements ritualisés, tenter d’inciter un éthologue à devenir un compagnon de vol, ou une femme de chambre, à devenir une partenaire sexuelle. Uexküll insiste sur le fait que la tonalité de l’Umwelt suffit : les tonalités de chasse ou de reproduction, projetées sur n’importe quelle figure de l’Umwelt, déclenchent l’action du choucas : incitation d’envol, ou parade nuptiale, en dépit de ce qui est perçu, et qui devrait être dissuasif.
- Note de bas de page 56 :
-
C’est précisément ce qui conduira Uexküll à envisager les « signes magiques » (des signes de quelque chose qui n’existe pas — pas encore — ou qui est absent). C’est le cas du crapaud affamé qui dévore un ver de terre et se précipite ensuite sur une allumette (p. 140).
Ce type de comportements repose sur les propriétés des tonalités, et plus particulièrement sur la plasticité des scénarios thématiques qui leur procurent une structure actantielle : certaines positions actantielles requises par la pratique peuvent rester en effet provisoirement vides, en attente d’un éventuel remplissement. La tonalité peut alors donner lieu à une prévision, qu’Uexküll appelle une image-prospection (pp. 137-143) : une tonalité spécifique étant convoquée, ou même déjà actualisée, l’animal peut percevoir autre chose que ce qui est effectivement présent dans son milieu, et la tonalité peut donc faire mentir la perception56. Remplir une place vide par un acteur qui n’est pas prévu dans les schèmes de l’espèce, sous la pression modale et affective d’une tonalité, et ce au prix d’un changement de repères véridictoires, implique parfois un changement de mode d’existence.
Un mode d’existence est principalement caractérisé en effet par un référentiel de véridiction (ce qui peut être « vrai » ou « mensonger » dans l’un peut ne pas l’être dans un autre) et par des règles de constitution du collectif en interaction. Au moins du point de vue anthropomorphe de l’éthologue, ces référentiels et ces règles participent, pour un même mode d’existence, au régime de croyances et aux conditions d’efficience et d’existence de ce dernier. Lorsque par exemple l’animal adopte un partenaire d’une autre espèce pour l’appeler à jouer un rôle dans son propre Umwelt et dans sa propre tonalité, le mode sous lequel il le propose aura pour le partenaire en question un caractère semi-fictionnel, voire ludique. Imaginons que l’autre réponde à son appel : ce sera une sorte de jeu, dont les conséquences resteront circonscrites à l’espace-temps du jeu, sur le même principe que la fiction.
Certes, pour l’animal qui le propose, il s’agit seulement d’une extension interspécifique dans la mise en œuvre d’une tonalité et de la prégnance biologique sous-jacente. Mais l’autre, en acceptant la proposition, valide cette extension et entérine une interaction qui se déroule alors sous un régime ludico-fictionnel. Pour mieux cerner cette substitution de mode d’existence, on peut comparer ce type d’interactions, entre deux êtres vivants, avec les interactions avec des objets inanimés. Si un crapaud affamé (tel que l’évoque Uexküll dans la présentation d’une expérience, p. 40) se jette sur un bout de bois pour le dévorer, aucune frontière entre modes d’existence n’est franchie : la pression de la prégnance biologique a été suffisante pour provoquer une extension du domaine d’application de la tonalité d’alimentation mais on en reste à une extension erronée, à l’intérieur du mode d’existence d’origine. Erronée, parce que le crapaud ne se nourrit pas de bouts de bois. Ce mode d’existence, où les prégnances biologiques peuvent se réaliser au bénéfice de la persistance de l’individu ou de l’espèce, peut être qualifié de vital. En revanche, si l’extension sollicite un partenaire animé et vivant, un autre animal, et même si les prégnances biologiques ne peuvent se réaliser physiquement que sous un régime strictement vital, l’Umwelt bascule dans une sorte d’« imaginaire », un monde où l’interaction peut se poursuivre sous un autre régime.
Ce régime ludico-fictif apporte une confirmation essentielle au rôle de la sensibilité dans l’autoconservation. En effet, dans un cas comme dans l’autre, la prégnance biologique a contribué à la mise en œuvre d’une tonalité, et nous savons que la réalisation des objectifs pratiques de cette tonalité, quelle qu’elle soit (alimentation, reproduction, défense et attaque, etc.) contribue à l’autoconservation. Nous devons ajouter alors : elle y contribue, quel que soit le régime sémiotique et le mode d’existence où elle se réalise. Dans le mode « vital », cela va de soi. Mais dans le mode « ludico-fictif », la tonalité se réalise aussi, mais principalement dans sa dimension sensible et réflexive (cf. supra, à propos du centre réflexif de sensibilité-activité).
Il semblerait bien que nous soyons conduits ici à imaginer, tout comme les anthropologues contemporains pour les collectifs homme-nature, un double fonctionnement des entités vivantes, en extériorité pour les aspects physiques, et en intériorité pour les aspects sensibles et réflexifs : pour le choucas qui cherche à séduire une femme de chambre, il n’y a aucun espoir d’assurer ainsi la survivance physique de son espèce (question anthropomorphe : « En doute-t-il ? »), mais il n’en assure pas moins le renforcement et la survivance de la dimension sensible et réflexive de son propre Umwelt, avec la contribution bien involontaire de la femme de chambre. Or l’Umwelt est à la fois un ensemble d’interactions indivisible, et une condition requise pour la persistance d’une espèce : la persistance de la sensibilité réflexive est à cet égard tout aussi efficiente que la satisfaction des besoins physiques vitaux.
- Note de bas de page 57 :
-
Dans son chapitre consacré aux milieux magiques, Uexküll accorde en effet beaucoup de poids à ce qu’il appelle la « subjectivité » : les acteurs imaginaires inventés par l’animal, ainsi que les projections de rôles « erronés » ou irréalistes qu’il accomplit à l’égard des acteurs inadaptés aux tonalités en cours, sont pour l’éthologue des manifestations de la subjectivité animale. Mais, même euphémisée, comme nous le proposons, en « subjectalité », cette explication reste insatisfaisante : ce sont au contraire les caractéristiques quasi-énonciatives propres aux tonalités, et notamment celle qui leur permet de passer d’un mode d’existence à un autre, qui suscitent ici l’effet de sens « subjectalité ».
Uexküll évoque dans ce cas les « milieux magiques », en faisant ainsi porter la pression tonale sur l’Umwelt tout entier. Il en tire argument pour conforter l’idée que les mondes animaux sont « subjectifs ». Sans prêter à l’animal une subjectivité, une capacité d’imagination, ou des croyances magiques, on peut au moins lui reconnaître une propension à changer de mode d’existence au cours des interactions, et à manifester ainsi le double fonctionnement (indivisible) de son Umwelt, à la fois sur la dimension des extériorités physiques et sur la dimension des intériorités sensibles et réflexives57.
Sensibilité aux circonstances des interactions et à la « deixis » fixée par le centre de sensibilité-activité de l’Umwelt, sélections parmi plusieurs alternatives, embrayages et débrayages entre niveaux et modes d’existence : ce sont autant de caractéristiques et de potentiels dynamiques des tonalités qui concourent à les rapprocher de l’énonciation, entendue comme acte d’instauration et d’actualisation de mondes vivants, et comme ajustement pratique des schèmes disponibles à la situation particulière où ils sont actualisés. Dans cette perspective, c’est la pression tonale (thématique et affective) qui fournit le régime de signification de l’interaction globale, qui garantit les liaisons internes de la pratique, et qui produit des sémioses intégrales et interprétables.
3.3.3. Une sémiotique modulaire et énonçable
Uexküll utilise les termes stimulus, signal, signe, et image. Nous avons déjà constaté que cette segmentation, qui présente l’avantage de permettre d’identifier des unités du plan de l’expression, ne donne pas pour autant accès à la signification des interactions au sein de l’Umwelt. Par conséquent, si nous maintenons l’hypothèse selon laquelle les tonalités et les pressions qu’elles exercent sont les configurations et les forces qui permettent d’actualiser des schèmes et d’instaurer des mondes animaux, nous devons, comme pour n’importe quel autre type d’énonciation, préciser ce à partir de quoi elles énoncent et instaurent.
La réponse à cette demande se trouve dans les propriétés accumulées pour constituer les Umwelten, que l’on peut maintenant répartir en une série de modules méthodologiques :
- Note de bas de page 58 :
-
Les types de champs sensibles ont été décrits d’un point de vue sémiotique notamment dans J. Fontanille, Corps et sens, Paris, PUF, 2011.
i) Les modes du sensible : ce sont les modes tactile, olfactif, visuel, moteur, auditif, sensori-moteur, proprioceptif, etc. Chaque mode peut être décrit selon les propriétés du champ sensible (forme globale du champ, profondeur et orientation de la profondeur, type de relation entre actants sensibles, etc.)58.
ii) Les gammes sensorielles : des gammes pertinentes, plus ou moins étendues, sont sélectionnées dans chaque mode du sensible (par exemple un seul son très aigu, qui a les mêmes caractéristiques que le cri de la chauve-souris, peut alerter le papillon de nuit). Le champ sensible peut être ainsi caractérisé en extension (restreint / étendu).
- Note de bas de page 59 :
-
Certains animaux semblent, du point de vue humain, capables de reconnaître des formes ; par exemple le ver de terre serait capable de faire la différence entre la pointe et la tige d’une feuille d’arbre lorsqu’il l’entraîne dans son trou ; mais des expériences ont prouvé qu’il est « seulement » capable de reconnaître le goût différent des deux extrémités. En revanche, l’abeille reconnaît vraiment la forme ouverte et découpée de la fleur épanouie (même quand ce n’est pas une fleur), et la distingue de la fleur en bouton (même quand c’est vraiment une fleur). Il en résulte que l’identification des traits figuraux (la différence de goût plutôt que l’identification d’une orientation de la forme) est décisive dans la caractérisation et la spéciation de l’Umwelt.
- Note de bas de page 60 :
-
Le même objet ou le même « autre » peut être identifié de manière séparée, voire antagoniste par le même individu ; c’est le cas de la poule qui a couvé un poussin noir parmi ses poussins jaunes, et qui, d’un côté, vient à son secours et combat du bec un éventuel danger quand le poussin noir envoie un cri de détresse (signe auditif), mais de l’autre le chasse systématiquement quand elle le voit parmi les autres poussins (signe visuel chromatique). Les paramètres d’identification de l’autre ne sont donc pas systématiquement coordonnés, et peuvent donner lieu à des interactions divergentes. On sait également qu’il ne s’agit pas à proprement parler de deux Umwelten différents, mais de deux colorations tonales différentes (menace et secours quand la poule entend le poussin crier ; cohésion du « nous » et des mêmes quand elle ne voit que sa couleur noire).
iii) Les propriétés sensibles figurales : c’est le degré ultime de la co-sélection, quand elle ne retient que des figures plastiques ou génériques (le mouvement, ou la forme, ou la couleur, etc.), sans prendre en considération la figure iconique globale59. Ces types figuraux sont extraits des entités figuratives de l’environnement60.
- Note de bas de page 61 :
-
Pour compléter et développer ces régimes spatiaux, on peut se reporter à E. Landowski, « Régimes d’espace », Actes Sémiotiques, 112, 2010.
iv) Les régimes spatiaux : l’Umwelt est un lieu qui est analysé par Uexküll en plusieurs catégories distinctes : en endroits (localisation), en mosaïque d’endroits (plus ou moins dense et complexe), en chemins (direction), en repères (orientation), en lointains et proches (profondeur). Ces régimes spatiaux reposent sur des schèmes de parcours (les « chemins ») et de relations positionnelles (la demeure, la zone familière, comprenant la zone neutre et le territoire). Du point de vue topologique, les distinctions sémiotiques (topique pour la demeure, paratopique pour la zone familière, utopique pour l’ensemble du territoire, hétérotopique pour tout ce qui est hors Umwelt, et atopique pour la zone frontière neutre) devraient être complétées par des distributions modales de pouvoir et devoir faire sur chacun de ces types spatiaux61.
- Note de bas de page 62 :
-
Uexküll donne l’exemple de l’escargot qui est arrêté dans sa progression par un bâton qui bat deux ou trois fois par seconde, parce qu’il le prend pour un bâton en mouvement, mais qui tente de passer par-dessus un bâton qui bat au moins 5 ou 6 fois par seconde, parce qu’il le prend pour un bâton immobile.
v) Les régimes temporels : l’Umwelt se caractérise en premier lieu par la structure de l’instant, qu’Uexküll décrit a minima et entre autres par une fréquence rythmique (le nombre de battements par instant62), ou par des étirements et des suspens de l’instant. Cette structure de l’instant peut avoir des corrélats affectifs, comme l’attente, ou du moins l’affect du « temps suspendu » qui en tient lieu chez les animaux, comme la tique.
vi) Le nombre et la structure des cycles perception / action : les séquences pratiques canoniques de chaque espèce se caractérisent par (a) le nombre de cycles ; (b) le fait que ce sont des cycles autonomes et harmonisés, ou pas ; (c) l’existence de cycles externalisés dans un organe dédié, ou pas ; (d) le fait que ces cycles soient commandés par un régulateur unique, ou par plusieurs. La régulation de ces cycles fonctionnels implique des schèmes, repris dans des tonalités thématiques et existentielles qui assurent les transitions et les tensions entre les cycles successifs.
vii) Les schèmes : des schèmes se constituent par apprentissage, par mémorisation, par imitation, par transmission inter-groupale et/ou intergénérationnelle. Il faut distinguer le processus de schématisation (apprentissage, imitation ou transmission), et le type de disponibilité et de motivation (notamment tonale) du schème en vue de sa réplication (mémoire, génétique, culture d’espèce ou de groupe).
Les tonalités, à la fois actionnelles et affectives, projettent une « coloration » modale et thématique sur l’Umwelt, qui actualise tous les éléments compatibles des sept modules précédents, pour en faire un monde cohérent, ici et maintenant. Uexküll ébauche une typologie des tonalités très hétérogène, le plus souvent ad hoc : tonalités d’alimentation, de nutrition, d’amour, de défense (donc des prégnances et des schèmes pratiques de base), mais aussi tonalités d’obstacle, d’ascension, d’éclairage, de désaltération, de s’asseoir, de déambulation, etc. Une typologie raisonnée et hiérarchisée des tonalités est donc encore à construire.
3.3.4. Eléments de méthode
Du point de vue de l’analyste, qui vise une caractérisation singulière de chaque monde vivant actualisé sous une tonalité donnée, la description du régime sémiotique de chacun d’eux pourra notamment prendre en considération :
1) le taux de sélectivité et de dissociation des propriétés sensibles figurales,
2) les régimes spatiaux et temporels,
3) les types de schèmes syntagmatiques : improvisés, programmés, adaptés, etc.,
4) la complétude ou l’incomplétude des scènes prédicatives convoquées par les tonalités (cf. les cas où un rôle prévu n’est attribué à aucun acteur, ou à un acteur incompatible avec la pratique en cours),
5) le mode d’existence du « remplissement actoriel » des rôles actantiels thématiques,
6) le rôle et l’intensité des pressions affectives.
Pour accompagner cette démarche, nous sommes maintenant en mesure de proposer un tableau récapitulatif de l’ensemble de la sémiotique tonale d’Uexküll, en y distinguant trois dimensions complémentaires : i) les sélections sémantiques, ii) les organisations syntagmatiques, et iii) les tonalités existentielles.
Soit une répartition classique entre sémantique, syntaxe et énonciation-instauration. Les modes d’instauration-énonciation procurés par les tonalités correspondent majoritairement à des prégnances biologiques (défense, reproduction, alimentation, etc.) ; les sélections sémantiques portent sur les manifestations figuratives des précédentes, qui correspondent à des saillances biologiques ; et les organisations syntagmatiques résultent de l’ensemble des processus de schématisation.
Les plans d’analyse, définis ci-dessus au point 3.3.3, sont disposés verticalement dans le tableau suivant, ce qui suggère une hiérarchie : si cette suggestion était érigée en hypothèse, et si cette hypothèse était vérifiée, l’ensemble pourrait alors constituer éventuellement un parcours génératif de la signification des mondes animaux.
|
Sémantique de l’Umwelt |
Syntaxe de l’Umwelt |
Instauration de l’Umwelt |
|
|
Plans d’analyse |
Sélections sémantiques |
Schèmes syntagmatiques |
Tonalités existentielles |
|
Perception figurative et figurale |
Modes du sensible Gammes sensorielles Types sensoriels figuraux |
Nombre et structure des cycles fonctionnels (perception/action) |
Tonalités perceptives de l’Umwelt |
|
Régimes spatiaux |
Lieux, endroits, repères, profondeurs |
Chemins et zones de familiarité, affects spatialisés |
Prégnances spatiales : (demeure, territoire) |
|
Régimes temporels |
Types temporels : structures de l’instant |
Schèmes temporels : rythmes et affects temporalisés |
Prégnances temporelles |
|
Actants et acteurs |
Modes d’identification des acteurs de l’Umwelt |
Rôles et modalisations des figures de l’Umwelt dans les interactions |
Tonalités comme modes d’existence des actants et acteurs |
|
Pratiques |
Prédicats et scènes prédicatives |
Schèmes et séquences pratiques de l’espèce |
Prégnances et schèmes de base |
Pour compléter ces propositions méthodologiques, nous pouvons enfin distinguer deux grandes dimensions pour la caractérisation des interactions, qui conduisent à un début de typologie des régimes sémiotiques des Umwelten. Ces deux dimensions sont d’une part les régimes sémiotiques de la consistance des interactions, qui concernent principalement les sélections sémantiques et les organisations syntagmatiques, et d’autre part ceux de leurs tonalités existentielles, qui concernent les modes d’énonciation-instauration, ainsi que les modes d’existence qui en découlent.
L’étude de la signification des Umwelten doit viser les particularités des espèces, des groupes vivants ou des individus, y compris dans des situations atypiques et singulières : la description vise la spécification, voire l’individuation des régimes de signification. Nous retrouvons ainsi les problématiques traitées par la socio-sémiotique des humains : chacun des différents types d’interactions procure des conditions pour un ou des régimes sémiotiques spécifiques.
Les conditions de spécification et d’individuation identifiées dans les analyses d’Uexküll ont déjà été inventoriées dans le tableau précédent : en chaque ligne et en chaque colonne, des sélections et des orientations sont opérées, et on obtient ainsi un nombre considérable de variables et de variétés. Pour parvenir aux typologies respectives de la consistance des interactions et des tonalités existentielles, il nous faut réduire cet inventaire hiérarchisé à un petit nombre de domaines déterminants pour les régimes sémiotiques. Nous retenons ainsi :
a) La sélectivité. Les discussions philosophiques sur les mondes animaux ont souvent porté sur la pauvreté ou la richesse des mondes sensibles respectifs des animaux et des êtres humains. Uexküll fait état de ces discussions mais leur oppose le fait que pour chaque animal la sélection de ce nous avons appelé des types sensibles, des gammes sensorielles et des types figuraux lui permet de distinguer le monde de son Umwelt, où il peut opérer en toute « certitude », et l’outre-monde, où tout devient incertain et incompréhensible. — La dimension de la sélectivité concerne, par rapport au tableau qui précède, la colonne intitulée « sélectivité sémantique » (plus ou moins restrictive et radicale) ;
b) Les contraintes syntagmatiques. Cette deuxième dimension caractérise la nature et la force des pressions qui s’exercent sur les enchaînements de comportements. Selon le degré d’intensité de la contrainte, le comportement de l’animal sera plus ou moins prévisible, plus ou moins dicté par des réflexes, plus ou moins éloigné du fonctionnement d’une « machine » cartésienne. Les contraintes peuvent être accidentelles, occasionnelles, récurrentes ou systématiques. — La dimension des contraintes syntagmatiques porte, dans le tableau précédent, sur la colonne intitulée « schèmes syntagmatiques » ;
c) La diversité et l’hétérogénéité des modes d’existence disponibles. Pour un Umwelt et un animal donnés, nous avons vu qu’on peut aller et venir entre deux ou plusieurs modes d’existence (notamment eu égard à leur référentiel de véridiction et à leurs « régimes de croyance » ou le régime d’efficience). Ces possibilités sont variables selon les espèces et les individus : par exemple, les « erreurs » du choucas, qui cherche à impliquer un éthologue ou une femme de chambre dans ses pratiques de base, s’expliquent en partie par leur présence familière dans son environnement ; tous les choucas, évidemment, ne fréquentent pas assidument les éthologues et les femmes de chambre. De même, pour des raisons de degré de complexité de l’organisme et de variété de son milieu, l’Umwelt du ver de terre et celui du chien n’offrent pas les mêmes latitudes de changement de modes d’existence. — Cette troisième dimension n’apparaît pas dans le tableau précédent car il a été constitué uniquement à partir des catégories qu’on peut directement reconstituer à partir de la « sémiotique tonale » d’Uexküll, alors que l’introduction des « modes d’existence » est entièrement de notre fait ;
d) La force des pressions tonales. Si nous avons insisté sur la correspondance entre les tonalités et les prégnances biologiques, c’est principalement parce que même si on ne peut pas confondre une configuration sémiotique et une donnée biologique, la première doit au moins hériter de toutes les propriétés symboliques de la seconde. Et tout particulièrement, l’intensité d’une pression biologique concerne directement la sensibilité de l’organisme vivant sur laquelle elle s’exerce, et cette intensité se trouve donc nécessaire dans une configuration qui accorde une place centrale aux interactions sensibles. — Cette quatrième dimension correspond, dans le tableau ci-dessus, à la colonne « Tonalités existentielles ».
Ces quatre dimensions caractérisent le fonctionnement des régimes sémiotiques des Umwelten de manière graduelle, soit en intensité (les contraintes syntagmatiques et les pressions tonales), soit en extensité (la sélectivité, et la disponibilité des modes d’existence dans les Umwelten).
Les deux premières dimensions, la sélectivité et la force des contraintes syntagmatiques, définissent l’Umwelt caractéristique d’une espèce en général, quelles que soient les conditions concrètes et les circonstances dans lesquelles il se réalise et donne lieu à des occurrences particulières ou individuelles. La sélectivité est héritée à l’intérieur de l’espèce, notamment en raison des arcs réflexes qui associent les stimuli reçus du milieu et les actions et réactions de l’organisme. Les contraintes syntagmatiques sont elles aussi imitées, transmises, et héritées. On examinera donc d’abord et séparément, au titre de la consistance des interactions, la corrélation entre la sélectivité sémantique (les saillances) et les contraintes syntagmatiques (les modes de schématisation).
La diversité et la disponibilité des modes d’existence, et les variations d’intensité des pressions tonales, en revanche, déterminent directement la manière dont les Umwelten sont réalisés, sous forme d’occurrences singulières, car toutes deux concourent à l’appropriation et à l’adaptation de l’Umwelt générique de chaque espèce par les individus, et selon les circonstances. On examinera donc ensuite, au titre des tonalités existentielles, la corrélation entre la force des pressions tonales (issues des prégnances) et la diversité des modes d’existence disponibles (la composition existentielle variable des scènes prédicatives).
1) Régimes sémiotiques de la consistance des interactions dans les Umwelten
- Note de bas de page 63 :
-
On aura reconnu l’écho direct, dans le monde animal, de la typologie des régimes de sens dans l’interaction établie par Landowski (Les interactions risquées, op. cit.).
Les interactions entre entités vivantes, et entre organismes et milieu, sont plus ou moins rigides, plus ou moins prévisibles, et plus ou moins contrôlées ; elles peuvent être entièrement programmées, ou adaptées, voire inventées pour faire face à l’occasion ou aux aléas63. C’est ce que nous appelons les « régimes de consistance des interactions », qui gèrent plus précisément la manière dont la relation entre l’organisme et son milieu est constituée en Umwelt générique :
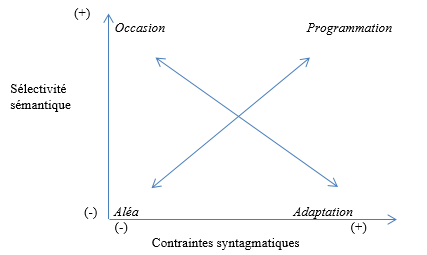
L’occasion et l’aléa se caractérisent tous deux ici par une faible contrainte syntagmatique, mais se différencient par le fait que la première, contrairement au deuxième, l’associe à une forte sélectivité sémantique. De même, la programmation et l’adaptation sont toutes deux sous une forte contrainte syntagmatique, mais se différencient par le fait que la première, contrairement à la deuxième, l’associe à une forte sélectivité sémantique. Le régime de l’aléa est le moins déterminé, et comme le rappelle fréquemment Eric Landowski, la seule question qui se pose à son égard est de savoir si on le prend en considération comme contingent, si on l’évite comme malencontreux, ou si on l’intègre comme opportunité. L’occasion n’échappe que partiellement à la contrainte syntagmatique, dans la mesure où elle peut advenir de la conjoncture entre plusieurs contraintes indépendantes. Mais pour que l’organisme vivant puisse se saisir d’une telle conjoncture, il faut que les dimensions de la sensibilité requises soient effectivement déjà héritées et activées. La différence entre la programmation et l’adaptation est de même nature : sur le fond de fortes contraintes syntagmatiques, la programmation est mise en œuvre à partir de sélections sémantiques déjà héritées et actives, alors que l’adaptation consiste justement à opérer des sélections qui ne sont pas encore en place.
C’est pourquoi, du point de vue des transformations, l’adaptation, par exemple, peut devenir programmation grâce à l’augmentation de la sélectivité sémantique (qui restreint au minimum les marges de l’adaptation) ; mais elle peut devenir aussi occasion, si les contraintes syntagmatiques s’abaissent, en même temps que la sélectivité augmente ; elle peut enfin se réduire à un pur aléa, dès lors que les seules contraintes syntagmatiques s’affaiblissent.
2) Régimes sémiotiques de la tonalité existentielle des interactions
Les prégnances sont prises en charge plus ou moins intensément par les pressions tonales, plus ou moins directement ou indirectement dans les modes d’existence disponibles. Ces modes d’existence disponibles peuvent être réduits à un seul ou nombreux. Ces deux variables engendrent la diversité des régimes de la tonalité existentielle.
La terminologie retenue découle directement des analyses d’Uexküll, et des effets de sens induits par les pressions tonales et les prégnances biologiques. On voit bien ici que les régimes ludique et imaginaire, à la différence des deux autres, exploitent autant que possible la disponibilité de plusieurs modes d’existence pour constituer la scène prédicative, et satisfaire les prégnances par tous les moyens disponibles. En outre, les régimes vital et imaginaire, à la différence des deux autres, sont ceux qui sont le plus fortement soumis à la pression tonale des prégnances, et ils diffèrent l’un de l’autre en raison de la diversité des modes d’existence : un seul pour le régime vital, une multitude pour le régime imaginaire.
Le régime accidentel est le moins déterminé : ni la pression tonale, ni la disponibilité des modes d’existence ne l’expliquent. Il permet au moins de prévoir, dans la typologie, une situation existentielle qui n’obéit à aucune détermination habituelle, du moins sous les conditions fixées par l’Umwelt. Cela ne signifie donc pas que l’accident est inexplicable ou ininterprétable, mais seulement qu’il échappe aux règles actuelles de l’Umwelt. Il faut rappeler ici que ce sont justement des accidents dans les mondes animaux, y compris et même surtout au niveau d’analyse génétique, qui contribuent, grâce à la sélection qui les retient ou les ignore, aux bouleversements des espèces et de leurs Umwelten.
Dans nos études de cas (supra, 3.3.2), nous n’avons pas clairement distingué le jeu et l’imaginaire, parce que nous ne disposions pas alors des critères adéquats pour le faire. Les deux régimes, le ludique et l’imaginaire, exploitent les possibilités de changement de mode d’existence, mais avec des enjeux tout différents. Dans le régime ludique, l’animal est faiblement déterminé par les prégnances biologiques, et il peut donc aller et venir entre des tonalités différentes, emprunter provisoirement un schème syntagmatique imprévu, et tout cela sans conséquences vitales. Il exploite les marges de liberté que lui offrent les transitions entre les référentiels de véridiction : il peut ainsi simuler et dissimuler, détourner, voire mentir. En revanche, quand il entre dans le régime imaginaire, il est sous la pression d’une prégnance, sous une pression tonale qui vise une satisfaction « à tout prix ». Dans ce cas, il ne simule ni ne ment, il poursuit la réalisation d’une tonalité dans un autre régime que physique et vital. L’oiseau qui chasse sa femelle du nid pour inviter le directeur du zoo à prendre sa place (cf. supra) ne simule rien et ne joue pas : c’est une invitation sérieuse, par laquelle il réalise sur la dimension sensible la tonalité d’attachement « sexué ».
Les transformations peuvent être aisément explicitées : par exemple, en partant du régime accidentel, trois voies sont ouvertes : vers le régime ludique, grâce à un nouvel assemblage de modes d’existence ; vers le régime vital, grâce à une augmentation de la pression tonale ; ou vers le régime imaginaire, grâce, à la fois, à un changement de mode d’existence et à une augmentation de la pression tonale des prégnances.
Conclusion
Le monde vivant est depuis longtemps l’apanage des sémiotiques du signe, et pour l’essentiel, du paradigme peircien. Loin de le regretter, il faut au contraire valoriser cet effort continu de tous les chercheurs qui ont consacré leurs travaux au domaine biosémiotique pendant plusieurs décennies. Mais aujourd’hui, la prise de conscience écologique, l’attention croissante pour la condition animale, les transformations profondes de la réflexion anthropologique, nous créent à tous une obligation, qui est autant éthique et politique que scientifique. Cette obligation a pour première conséquence qu’aucun paradigme théorique et méthodologique ne peut s’exonérer d’apporter sa contribution à l’effort collectif. Le relatif silence des sémiotiques structurales sur ces questions a longtemps prévalu, mais depuis quelques années, de nombreuses voix s’élèvent, qui rompent ce silence devenu incompréhensible. Quelques-unes de ces voix ont été évoquées au fil de la réflexion qui précède.
Nous avons avancé, en introduction, l’hypothèse selon laquelle le silence relatif des sémiotiques structurales, pendant près de quarante ans, sur les questions touchant au monde vivant, pouvait s’expliquer par son attachement à une vision naturaliste, dont résultait notamment le grand partage entre les sciences de la nature et celles de la culture, la sémiotique n’étant supposée participer que de ces dernières. L’explication apparaît maintenant tout à fait insuffisante, au moins pour deux raisons.
Déjà, dans les années soixante, la sémiotique structurale inspirée par l’œuvre de Greimas était fortement étayée par sa référence à l’anthropologie structurale de Lévi-Strauss. Or, même un lecteur lambda, mais suffisamment attentif, pouvait remarquer à quel point Lévi-Strauss entremêlait dans ses descriptions et dans ses élaborations théoriques l’ensemble des composants des mondes vivants (animaux, végétaux, autant qu’humains), et même, bien au-delà, toute une cosmologie. Certes, ces mondes vivants et ces cosmologies n’étaient appréhendés principalement qu’à travers des productions sémiotiques et symboliques humaines, des mythes, des rituels, notamment. Mais pas seulement : il n’est que de constater avec quel soin l’anthropologue confrontait plusieurs types de sources, notamment documentaires, touchant aux mondes animaux et végétaux, pour étayer ses analyses des productions culturelles. Mais, quoi qu’il en soit, même si le point de vue restait globalement culturel et anthropomorphe, l’entier de la nature et des mondes vivants étaient convoqués, en arrière-plan ou au premier plan, selon le cas, des structures de l’imaginaire humain.
En outre, et surtout, l’essentiel de notre réflexion consiste à montrer en quoi l’organon principal, sinon exclusif, des sémiotiques structurales, était peu approprié à une approche pertinente des mondes vivants en général, et en quoi il se prêtait à des projections anthropomorphes intempestives : le cours tout entier de la discussion portant sur les biosémiotiques peirciennes est de fait le support d’un examen critique de l’appareil théorique et méthodologique des sémiotiques structurales, et notamment de leur approche de la narrativité. Et en même temps, il s’agissait de proposer des aménagements, des inflexions, des révisions, toutes fondées pourtant sur un potentiel d’évolution déjà impliqué dans l’organon sémiotique. Notre étude consacrée à la sémiotique de Jacob von Uexküll est emblématique à cet égard, puisque nous avons pu montrer que, parti de la sémiotique du signe (des signes) qui avait cours de son temps, l’éthosémioticien a progressivement déplacé la focale et mis l’accent sur ce que nous avons décrit comme des « pressions tonales thématico-affectives » et globales, qu’il appelait lui-même des « tonalités ». C’est l’ethosémioticien lui-même, et non son commentateur contemporain, qui substitue peu à peu une « sémiotique des ensembles signifiants » à une « sémiotique des unités signifiantes ». Avec ces « ensembles signifiants » que sont les Umvelten, nous sommes bien entendu fort loin d’une sémiotique de la textualité, mais l’homologie reste frappante et suggestive : les signes perceptifs et actionnels sont aux tonalités ce que les signes linguistiques ou visuels sont aux textes. Et une autre homologie est également apparue : les tonalités des interactions et les pressions tonales sont aux schèmes de comportement réflexe ce que les énonciations sont aux énoncés textuels.
En discutant des choix épistémologiques et méthodologiques des sémiotiques du signe, en matière biosémiotique, nous avions pour objectif — du moins est-ce ce que nous avons fait, quel que fût l’objectif d’origine — de nous approprier leurs résultats, et les raisonnements qui y conduisent, pour dégager les grands ensembles conceptuels et les principes méthodologiques susceptibles de conforter une démarche structurale. Ces ensembles conceptuels et ces principes méthodologiques — la conversion des explications téléologiques en conditions de validité narratives, la lecture rétrojective, la prise en compte globale des Umvelten comme sémiotiques-objets, la primauté des interactions et de leurs régimes sémiotiques sur toutes les autres élaborations secondaires, la place de la sensibilité et de la subjectalité, etc. — ne forment pas encore une théorie cohérente et complète, mais seulement son ébauche. Le chantier est ouvert, et bien entendu, à prolonger et à partager.