Diversité et paléosynécologie fonctionnelle de la paléoflore du bassin houiller de Bosmoreau-les-Mines (Creuse, FRANCE) Diversity and functionnal palaeosynecology of the palaeoflora from the Bosmoreau-les-Mines coal basin (Creuse, FRANCE)
David DELMAIL
et Pascal LABROUSSE
Un inventaire de la paléomacroflore stéphanienne a été réalisé dans le bassin houiller de Bosmoreau-les-Mines (Creuse, France) à partir d’empreintes issus des schistes. Des indications sur l’évolution des paramètres abiotiques, la paléobiologie et la paléosynécologie fonctionnelle permettent de mieux appréhender le fonctionnement du paléophytoécosystème. Le paléoenvironnement est dominé majoritairement par les Spermatophyta et notamment par les Cordaitales permettant ainsi de mettre en évidence (1) la plus ancienne réorganisation de forêts marécageuses du Carbonifère en écosystèmes fluviatiles complexes dans le Massif Central au Stéphanien et (2) le remplacement des Marattiales dominantes par les Cordaitales dans diverses niches écologiques. Toutes ces données établissent les bases d’études ultérieures de paléophytosociologie.
An inventory of the Stephanian palaeomacroflora has been drawed up in the coal basin of Bosmoreau-les-Mines (Creuse, France) from schist plant marks. Some indications on the abiotic-parameter evolution, the palaeobiology and the functional palaeosynecology allow a best understanding of the palaeophytoecosystem functioning. The palaeoenvironment is mainly dominated by Spermatophyta and especially Cordaitales which highlights (1) the earliest complete reorganization of Carboniferous swamp forests into complex fluviatil ecosystems in the Massif Central during the Stephanian and (2) the replacement of dominant Marattiales by Cordaitales in several ecological niches. All present data establish the fundaments of further studies about palaeophytosociology
Introduction
Bosmoreau-les-Mines est une commune située dans le département de la Creuse essentiellement connue pour ses anciennes exploitations houillères de 1784 à 1958. Suite à cette activité industrielle, de nouveaux paysages miniers se sont dessinés avec le déboisement et des monticules de terrils. Les terrils ont permis de mettre en évidence la présence de fossiles végétaux depuis 1868 (Gruner, 1868). Cependant, le bassin de Bosmoreau-les-Mines a suscité peu d’études scientifiques (Becq-Giraudon, 1985 ; Gruner, 1868) en comparaison des bassins contemporains comme celui de Montceau-les-Mines où la découverte de riches faunes a fait l’objet d’un regain de recherches (Poplin & Heyler, 1994) ou celui de Graissessac à proximité de l’institut de paléobotanique de Montpellier (Galtier et al., 1997). Aussi il nous paraissait essentiel de compléter le peu de données préexistantes en étudiant davantage la diversité de la paléomacroflore du bassin de Bosmoreau-les-Mines lors du stage « Introduction à l’étude des paléoenvironnements » de la SULIM (Station Universitaire du Limousin) du 18 au 21 Juillet 2009.
Comme dans tous les bassins houillers du Massif Central, c’est le fonctionnement d’une faille synsédimentaire qui est à l’origine de l’affaissement du bassin. Aussi longtemps que la faille est demeurée active, la subsidence s’est poursuivie, compensée par l’apport continu de la sédimentation (Galtier, 1999). L’âge des schistes de Bosmoreau-les-Mines est estimé entre 303,9 et 299 Ma (Stéphanien) à la fin de la période Carbonifère (Becq-Giraudon 1985). Comme dans le cas de tous les bassins houillers du Massif Central, il s’agit d’un bassin limnique ou intramontagneux par opposition aux bassins dits paraliques ou côtiers (à l’exemple du bassin du Nord-Pas-de-Calais). Le Massif Central faisait alors partie de l’immense chaîne de montagnes hercyniennes longue de plusieurs milliers de kilomètres (Galtier, 1999).
D’un point de vue climatique, le Massif Central se trouvait au Stéphanien en position équatorial sous un climat tropical chaud et humide favorable à l’installation d’une flore à haute diversité taxonomique à l’instar des forêts équatoriales actuelles (Poplin & Heyler, 1994).
Inventaire paléofloristique
La connaissance des associations végétales repose sur l’étude des macroflores et des microflores fossiles. Les macrorestes végétaux les plus communs se présentent sous formes d’empreintes (ou compressions) dans les schistes. Les fossiles de ce type ont pu être récoltés dans les anciens terrils des mines de charbon de Bosmoreau-les-Mines (Fig. 1) et ont été déterminés d’après Cleal & Thomas (2009) et Emberger (1944).
Figure 1. Emplacements des terrils fossilifères stéphaniens étudiés (indiqués par les pastilles numérotées). Photographie aérienne d’après Google Earth 5.0.1.

Gruner a publié dès 1868 une liste d’une dizaine de taxa. Bien plus tard, Becq-Giraudon (1985) a publié une autre liste un peu plus diversifiée.
La macroflore comprend de nombreuses Ptéridophytes et des Gymnospermes. On peut donner la liste suivante des taxa les plus communs trouvés en compressions :
-
Lycophyta
-
Lépidodendrales : très peu de restes de Sigillaria (tronc), Cyperites (feuilles), Stigmaria (système racinaire) sont présents.
-
-
Sphenophyta
-
Calamitales : très abondantes, divers troncs de Calamites (Fig. 2A) avec des branches verticillées (Asterophyllites) et des feuilles (Annularia).
-
-
Filicophyta
-
Marattiales : présence notable et diversité de fougères arborescentes (Psaronius) avec frondes de Pecopteris.
-
Filicales primitives : des fougères de dimensions modestes, à frondes délicates de type Sphenopteris, sont bien plus rares.
-
-
Spermatophyta
-
Ptéridospermales : abondantes et très diversifiées avec deux ordres pour un total d’au moins six genres. Les frondes de Callistophytales les plus communes sont de type Alethopteris (Fig. 2B), Callipteridium, Dicksonites, Linopteris (Fig. 2C) et Odontopteris. Les Medullosales quant à elles sont représentées par des restes ovulifères de type Pachytesta.
-
Cordaitales : abondance de feuilles de type Poacordaites (Fig. 2D) et une faible fréquence de celles de type Cordaites.
-
Figure 2. Photographies des taxa majoritaires du bassin stéphanien de Bosmoreau-les-Mines : A, tronc cannelé de Calamites présentant une cicatrice axillaire (ca) (échelle = 5 cm) ; B, pinnules de Alethopteris (échelle = 1 cm) ; C, frondes de Linopteris (échelle = 5 cm) ; D, feuilles parallélinervées de Poacordaites (échelle = 3 cm).

Paléobiologie
La plupart des grands groupes présents comptaient des formes arborescentes correspondant à cinq catégories bien distinctes (Galtier, 1999).
-
Les Sigillaria pouvaient dépasser 20 mètres de haut pour un diamètre basal du tronc d’un mètre (Fig 3A). Ces plantes avaient une croissance rapide ; la forme juvénile de l’arbre était pourvue d’un tronc non ramifié presqu’entièrement couvert de longues feuilles. Le cylindre de bois qui représentait moins de 1/5 du diamètre total du tronc n’était pas suffisant pour assurer le support de si grands arbres. C’est une écorce secondaire fibreuse très développée qui assurait le rôle principal de soutien, ce qui correspond à un modèle biomécanique sans véritable équivalent dans la nature actuelle.
-
Chez les Calamites, la taille (jusqu’à 10 m de haut et 50 cm de diamètre) et le port étaient variables en fonction du degré de ramification du tronc (Fig 3B). Comme les prêles et les bambous, dont elles devaient avoir un peu l’allure, ces plantes étaient caractérisées par une moelle creuse, mais elles possédaient un cylindre de bois dense qui assurait le support du tronc. Après la mort de l’arbre, la moelle creuse se remplissait de sédiment et les fossiles les plus fréquents sont précisément des moulages internes de ces troncs.
-
Les Fougères arborescentes sont des Psaronius dont la tige pouvait atteindre 10 m de haut et les frondes (Pecopteris) 3 m de long (Fig 3C). Chez ces plantes dépourvues de bois, la stabilité du tronc était assurée par un manchon de racines adventives de plus en plus épais vers la base où il atteignait 1 m de diamètre. Ces racines masquaient la véritable surface de la tige, à l’exception du sommet du tronc où l’on voit les cicatrices de feuilles déjà tombées. La tige obconique étroite à la base (quelques centimètres de diamètre) n’aurait pu tenir debout sans le système de racines adventives qui formait un ensemble cohérent.
-
Les Ptéridospermales étaient caractérisées par leurs grandes feuilles (jusqu’à 4 m de long) très semblables à celles des fougères mais le plus souvent bifurquées (Fig 3D). Ces formes ligneuses étaient de taille et de ports différents. Les plus grêles (tiges de quelques centimètres de diamètre) étaient autoportantes ou lianescentes. D’autres étaient des arbustes. Les seules ayant véritablement la stature d’arbres étaient certaines Medullosaceae dont le tronc atteignait 50 cm de diamètre. Ces plantes avaient un port de Fougère arborescente mais elles possédaient du bois. Ce tissu en combinaison avec l’écorce externe sclérenchymateuse, assurait le support du tronc.
-
Les Cordaitales sont parfois considérées comme les seuls arbres authentiques, dans la mesure où leur tronc était constitué essentiellement d’un bois dense très semblable à celui des conifères actuels (Fig 3E). Leur taille pouvait dépasser 30 m pour un tronc de plus d’1 m de diamètre. Leurs feuilles rubanées, rigides, à nervures parallèles, avaient jusqu’à 1 m de long et 10 cm de large.
Figure 3. Illustrations des 5 grands groupes végétaux présents dans le bassin stéphanien de Bosmoreau-les-Mines : A, Sigillaria ; B, Calamites ; C, Psaronius ; D, Medullosa ; E, Cordaites.
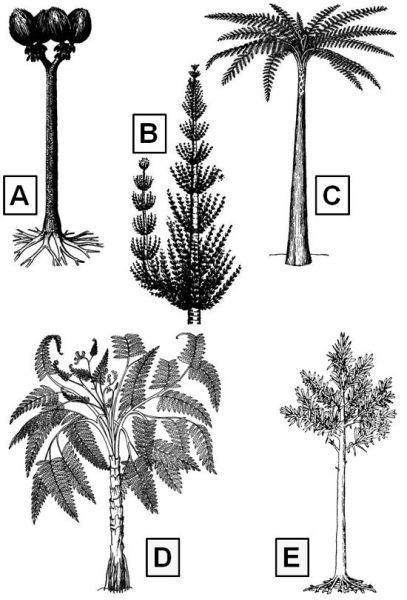
Dessins issus du site internet http://dic.academic.ru
Paléosynécologie fonctionnelle et évolution environnementale
Cette paléoflore recouvre la plupart des associations végétales contemporaines inféodées à diverses niches écologiques. D’un point de vue diversitaire, les taxa majoritaires étaient les Calamitales, les Cordaitales et les Ptéridospermales. Cette diversité au niveau des ordres contraste avec la diversité du Carbonifère moyen et inférieur notamment dans les bassins paraliques où la domination des Lycophyta arborescentes était avérée car elles étaient adaptées aux environnements marécageux nombreux durant cette période par le climat pseudo-tropical régnant sur la majorité des continents. Cependant dès le Carbonifère supérieur, le climat s’assèche et engendre la réduction progressive des marais, laissant progressivement la place aux Filicophyta et aux Spermatophyta plus aptes à croître dans les environnements dessicants. Les Lycophyta arborescentes, incapables de s’acclimater ni de s’adapter sur plusieurs générations aux changements abiotiques, ont vu leur diversité se réduire avec l’extinction de genres majeurs (e.g. Lepidodendron, Lepidphloios, Diaphorodendron et Synchysidendron) jusqu’au Permien inférieur (de 299 à 270,1 Ma) avec la disparition de leurs ultimes représentants du genre Sigillaria (Dimichele & Phillips, 1996 ; Galtier, 1999 ; Meyer-Berthaud com. pers., 2007).
Cependant, la diversité paléoflorale de Bosmoreau-les-Mines diffère nettement de celles des bassins contemporains du Massif Central car les Cordaitales sont ici bien plus dominantes que les Marattiales. En effectuant un relationnel avec les affinités écologiques de ces taxa, on constate que le bassin de Bosmoreau-les-Mines a certainement dû s’assécher plus rapidement que les autres bassins de la même région. En effet les Marattiales, bien que pouvant tolérer un environnement moins humide que les Lycophyta arborescentes, nécessitent des zones forestières légèrement marécageuses (Stidd, 1974). Cependant, les Cordaitales sont des végétaux de ripisylve (Raymond, 1988) pouvant tolérer un certain degré de dessication. Aussi il est fort possible que le bassin de Bosmoreau-les-Mines fut parmi les premiers bassins du Massif Central à subir les effets du changement climatique. Ses niches écologiques se sont restructurées totalement avec une réorganisation complète du système fluviatile causant une régénération des niches écologiques où la domination des Pteridophyta est estompée par celle des Spermatophyta.
Il est donc vraisemblable d’admettre que le bassin était une dépression d’altitude moyenne ou basse partiellement occupée par un lac et avec un réseau fluviatile complexe. La plaine alluviale était traversée par un fleuve alimenté par des torrents dévalant des montagnes hercyniennes. En raison du dépôt des matériels clastiques (i.e. issus de l’érosion), le remplissage du lac s’est effectué progressivement pendant quelques millions d’années et a permis de conserver les restes de paléoflore déposés en son fond. Dans les terrils de Bosmoreau-les-Mines, peu de cyclicité des couches de charbon est observée. Cela indique que le bassin durant cette période stéphanienne n’a pas été touché par des fluctuations cycliques du niveau des eaux confirmant que la faille synsédimentaire avait ralenti nettement son activité de subsidence et qu’un microclimat stable sans alternance d’épisodes secs et humides s’était mis en place.
Conclusion
Figure 4. Reconstruction paléoenvironnementale du bassin de Bosmoreau-les-Mines au Stéphanien.
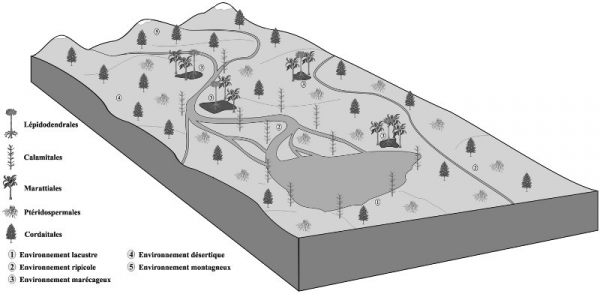
Le paléoenvironnement du bassin de Bosmoreau-les-Mines était complexe et composé de diverses niches écologiques (Fig. 4). Les différences écologiques étaient liées aux paramètres abiotiques tels les variations d’altitude, de température et d’hygrométrie. Les associations de végétaux plus ou moins xérophiles regroupaient les Spermatophyta qui se distribuaient dans la ripisylve ou les reliefs moins alimentés en eaux. D’un point de vue taphonomique (i.e. processus intervenant après la mort d'un organisme jusqu'à sa fossilisation), leurs restes végétaux étaient certainement dispersés par le vent vers les zones de rivière avant de se trouver véhiculés jusqu’à la zone lacustre où la sédimentation se réalisait. Les Ptéridospermales étant moins tolérantes à la dessiccation, elles étaient très probablement distribuées jusqu’en limite des zones désertiques avec à proximité une alimentation hydrique alors que les Cordaitales pouvaient s’y développer plus aisément ainsi que dans les environnements montagneux. La présence réduite d’associations de végétaux hélophytes (Filicophyta, Lépidodendrales) dont le développement se faisait le système racinaire dans des environnements hydriques anoxiques indique la réduction considérable des zones marécageuses. Une autre catégorie de taxa hélophytes regroupant uniquement les Sphenophyta, mais inféodés cette fois aux zones inondées ponctuellement, est fortement représenté en terme qualitatif et quantitatif. Aussi le pourtour de l’environnement lacustre où se développaient ces Pteridophyta devait être une zone très plane où l’eau pouvait rapidement inonder les berges sans qu’il y ait de grandes fluctuations du niveau moyen du lac comme évoqué précédemment.