Les peuplements d'Orthoptéroïdes de la lande sur serpentinite de la Flotte et du Cluzeau (Haute-Vienne)
Les peuplements d’Orthoptéroïdes d’une lande sur serpentinite du Limousin sont décrits pour la première fois. 24 espèces ont été dénombrées, représentant environ 37 % de la faune limousine. Le milieu le plus riche en espèce est la pelouse à Festuca lemanii. La lande sèche à Ajonc nain a un peuplement conforme à celui des autres landes comparables sur sol siliceux en Limousin. Les associations s’intègrent majoritairement dans les synusies euryméditerranéennes et eurosibériennes. Les successions bien caractérisées de peuplements que l’on observe associés à la dynamique végétale peuvent servir de guide pour la gestion de ces milieux soumis à la déprise agricole.
Orthopteroïd communities living on different plant associations settled on serpentinite formations are described for the first time. 24 species were recorded, representing about 37 % of the Limousin fauna. The richest milieu is the Festuca lemanii association. The dry heathland with dwarf gorse has the same community as in similar heathlands on siliceous soil in Limousin. Most insect communities belong to eurymediterranean and eurysiberian synusies. Well characterised successions of insect communities are closely associated to vegetation dynamics and constitute powerful tools to help in the management of these non-upkept habitats.
Introduction
Les formations végétales sur serpentinites sont parmi les plus originales du Limousin. Bien qu'il n'y ait pas de plantes endémiques, la rareté de certaines espèces, comme Festuca lemanii ou Notholaena maranthae, a été reconnue depuis longtemps. Ces particularités s'expliquent par un faisceau de conditions édaphiques et microclimatiques originales. En effet, les serpentinites sont des roches vertes issues d’anciennes péridotites pouvant former des chaos, dont l’altération conduit à des arènes ayant un fort pouvoir drainant, un pH proche de la neutralité et une composition chimique originale avec de nombreux cations, parmi lesquels le magnésium représente plus de 90 % du total. Les sols (Berre et al., 1974) sont foncés, riches en fer et métaux lourds (nickel, cobalt et chrome) mais pauvres en éléments minéraux essentiels (potassium, calcium). De part la couleur foncée de ces rochers et des sols, il y a accumulation de chaleur. La combinaison "milieu drainant-accumulation de chaleur" en fait un milieu xérophile et thermophile. La conséquence est la sélection d'un petit nombre d'espèces végétales ayant des propriétés physiologiques particulières, comme l'ont montré les travaux de Verger et al. (2001). Selon ces auteurs, certaines espèces assimilent peu ces cations et sont donc protégées naturellement de leurs effets toxiques, comme les fougères. D'autres, en revanche, comme F. lemanii, assimilent préférentiellement ces cations au niveau des racines et s’opposent à leur migration au niveau des feuilles, ce qui en limite les effets néfastes pour les consommateurs. On peut se poser la question de savoir si les plantes accumulatrices ne sont pas toxiques pour les insectes phytophages. Des expériences dans ce sens seraient nécessaires pour appréhender le sujet.
On peut s'attendre à ce que les peuplements orthoptérologiques présentent des traits originaux dans la mesure où leur composition reflète habituellement bien la structure végétale (exemples dans le Limousin : Picaud, 1998, Picaud et al., 2000). Nous avons donc entrepris d'analyser ces peuplements dans un gradient de dynamique végétale, depuis le rocher à peine colonisé jusqu'aux formations arborées. Les zones où le sol est peu épais montrent une pelouse à Festuca lemanii. De plus, sous l'effet de l'arrêt de la pression du pâturage extensif traditionnel depuis des dizaines d'années, on constate l'embroussaillement des landes à callune et ajonc nain avec un fort développement de la fougère aigle puis de la bourdaine et enfin des chênes sessiles.
Matériel et méthodes
Les milieux entourant le chaos rocheux de la Flotte et du Cluzeau sont à une altitude comprise entre 400 et 476 m, le point culminant étant situé à environ 4 km au SSW de Magnac-Bourg. Ils appartiennent aux communes de Château-Chervix, du côté de La Flotte, et de Meuzac, du côté du Cluzeau (Haute-Vienne). La carte (annexe 1) précise les emplacements des relevés.
Les milieux inventoriés suivent les groupements définis par Botineau (2000) :
-
Affleurements rocheux
Ordre de l’Asplenion serpentini (Asplenium adiantum-nigrum, Notholaena maranthae) -
Pelouse rase des ouvertures
Alliance du Sedo-Scleranthetalia (Sedum rupestre, Scleranthus perennis,..) -
Pelouse dense
Ordre du Koelerio-Phleion (Festuca lemanii, Thymus polytrichus subsp. britannicus, Armeria arenaria subsp. arenaria, Koeleria vallesiana…) -
Ourlet à Brachypode
Ordre du Trifolio-Teucrinenion scorodoniae (Filipendula vulgaris, Brachypodium pinnatum, Vincetoxicum arundinaria, Pteridium aquilinum,…) -
Variante de l'ourlet, pauvre en espèces, à Pteridium aquilinum
-
Lande sub-sèche
Association de l’Ericovagantis-Callunetum vulgaris (Ulex minor, Calluna vulgaris, Erica cinerea, Genista tinctoria) -
Lande mésophile
Association du Molinio-Ericetum vagantis (Erica tetralix, Erica ciliaris, Molinia coerulea,..) -
Boisement
Ordre du Frangulo alni-Pyrenion cordatae (Frangula alnus, Juniperus communis, Ilex aquifolium)
Sur des surfaces de 9 à 25 m² selon la saison (9 en Juillet et Août, 25 en Octobre), homogènes par la végétation, séparées de plus de 10 m les unes des autres pour éviter un effet de proximité, nous avons effectué 5 relevés exhaustifs dans plusieurs milieux différents. Le contour de chaque surface prélevée devait s'adapter aux limites des formations végétales : si dans la plupart des cas, il s'agissait de carrés, nous avons dû travailler parfois sur des bandes rectangulaires pour respecter l'homogénéité du groupement, ainsi que le préconise Guéguen (1976).
Les insectes ont été capturés en début (3/06/95), milieu (16/07/02) ou en fin de saison (28/09/02, 20/10/01) à l'aide de filets dans les zones herbeuses et de petits bocaux en verre dans les landes à Ajonc. Nous avons bénéficié dans cette opération de la coopération des étudiants de la maîtrise de Biologie des Populations et Ecosystèmes, mention Environnement de la Faculté des Sciences et Techniques de Limoges. Après détermination immédiate sur le terrain, les insectes sont relâchés sur place.
Les traitements statistiques ont nécessité plusieurs approches :
-
la richesse spécifique est le nombre d’espèces rencontrées.
-
l'indice de diversité de Shannon a été calculé grâce au programme SHAVA (Petit, 1998), selon la formule : H' = (ni/N log2 (ni/N), avec ni = nombre d'individus de l'espèce i et N = effectif total
-
les AFC (analyses factorielles des correspondances) et CAH (classifications ascendantes hiérarchiques) ont été effectuées grâce au logiciel GTABM (Quris, 1998).
-
les tests de Kruskal-Wallis et les calculs de corrélation de Pearson ont requis SYSTAT 7.0. (SPSS, 1997).
Résultats
1. Evolution saisonnière de la faune orthoptérologique
Nous présenterons l’évolution des paramètres de peuplement successivement par milieu et de manière globale.
1.1. Analyse par milieu
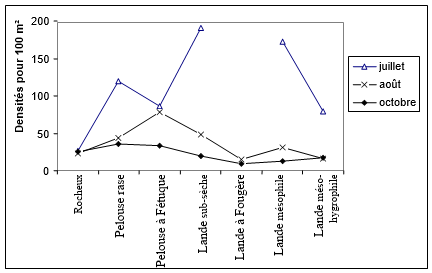
Figure 1. Evolution saisonnière de la densité par milieu
Les densités montrent une grande disparité saisonnière selon les milieux (fig. 1). Les affleurements rocheux, n'offrant que peu de ressources trophiques, sont peu peuplés. Dans la pelouse rase, la densité diminue d'environ de moitié (de 100 à 45) entre juillet et octobre. Dans la lande sub-sèche et dans la lande mésophile, elle est divisée par 8 environ de juillet à octobre (les chiffres d'août étant un peu supérieurs à ceux d'octobre).
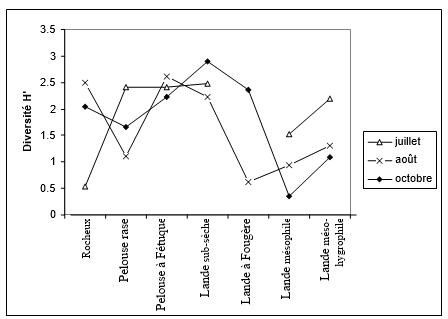
Figure 2. Evolution saisonnière de la diversité par milieu
La diversité H’ au niveau des affleurements rocheux est très faible en juillet par rapport au reste de la saison (fig. 2). En revanche, la pelouse rase offre en juillet une plus grande diversité que dans les mois suivants. Elle reste constante et de valeur élevée dans la pelouse à Fétuque, entre 2,3 et 2,6. La lande sub-sèche est également un milieu riche et plus particulièrement à la fin de la saison. Dans les landes colonisées par la fougère aigle, la diversité est très faible en août par rapport à celle d'octobre. Enfin, dans les landes mésophiles et mésohygrophiles, la diversité la plus élevée s'observe en juillet.
1.2. Analyse des paramètres de peuplement d’insectes
Les densités diminuent significativement de juillet à octobre (Kruskal-Wallis : N = 22, P = 0,006) puisque la moyenne, tous milieux confondus, est d'environ 110 pour 100 m² en juillet, 36 en août et 23 en Octobre. En revanche, le nombre d'espèces et la diversité de Shannon ne montrent pas de différences (Kruskal-Wallis : N = 22, P = 0,84 et P = 0,94 respectivement).
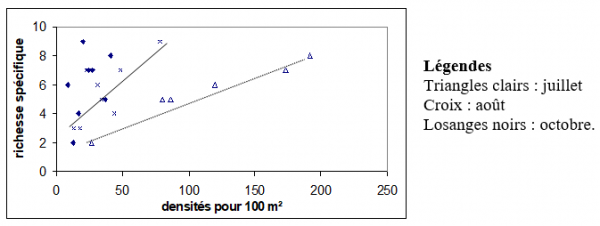
Figure 3. Nombre d’espèces et densité
En juillet (fig. 3), il y a proportionnalité entre la densité et la richesse spécifique (N = 6, r = 0,97, P = 0,001), relation qui est un peu moins marquée en août (N = 8, r = 0,76, P = 0,03) mais qui n'existe plus du tout en octobre (N = 8, r = 0,41, P = 0,34).
En ce qui concerne la diversité par rapport à la densité, aucune tendance ne se dégage pour chacun des mois étudiés (coefficients de corrélation non significatifs).
1.3. Analyse par espèce
Il y a un relais entre les espèces de début et de fin de saison, avec naturellement des espèces que l'on rencontre dans la durée. Les moyennes de densité des espèces, en confondant tous les milieux, sont rassemblées dans le tableau 1.
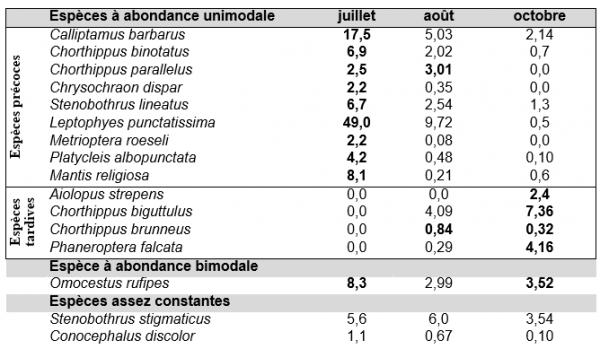
Tableau 1. Densités moyennes (pour 100 m²) des espèces fréquentes au cours de la saison. Certaines espèces rares ou mal échantillonnées ont été écartées.
On peut distinguer 9 espèces de début de saison, ou du moins dont les densités décroissent au cours de l'été, contre seulement 4 de fin de saison. Cette différence explique en partie les diminutions de densités au cours de l’été, particulièrement pour L. punctatissima. Le creux d'abondance d'Omocestus rufipes en août pourrait être en relation avec l’existence de 2 générations (la seule espèce de Caelifère dans ce cas sur ce site), comme dans le Midi de la France (Luquet, 1984). Cependant, certaines variations s'expliquent par la ± grande abondance d'une espèce selon l'année. Parmi les espèces non rapportées dans le tableau 1, Platycleis tessellata, par exemple, n’est pas particulièrement précoce en Limousin et n'a pas été vue en 2001 et 2002. L’ensemble des espèces recensées est indiqué dans l’annexe 2.
2. Les associations d'espèces
Nous avons effectué des AFC (d’après les tableaux des annexes 3, 4 et 5) séparément pour les 3 périodes d'étude, en raison des disparités de densité de la plupart des espèces. Les coordonnées des espèces sur les 3 premiers axes factoriels ont été considérées pour procéder aux calculs de distances car la somme cumulée des pourcentages de variance affectés à ces axes dépassait nettement 50 %. Les distances obtenues ont permis de produire les classifications hiérarchiques et à obtenir ainsi des regroupements objectifs d'espèces (fig. 4a, 4b et 4c).
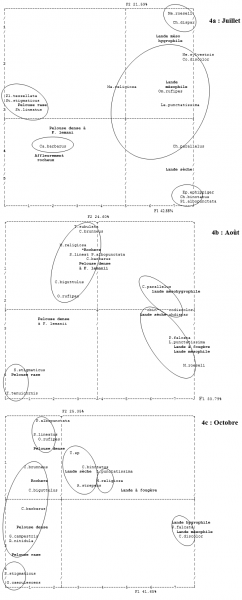
Figure 4. Projection des espèces et des milieux sur les 2 premiers axes de l'AFC
Les barres obliques indiquent l'emplacement de raccourcissements de figure, pour faciliter la mise en page.
Comment déduire les associations d'Insectes à partir de ces représentations ? Le problème vient du fait que les regroupements proposés par les statistiques présentent des points communs mais aussi des variations. Dans un souci descriptif d'établir une typologie des synusies du site étudié, il aurait été « agréable » d'obtenir les mêmes groupements tout au long de la saison, à l'exception des espèces soit précoces soit tardives.
Pour le raisonnement, nous choisissons de considérer la période intermédiaire comme base de référence, en raison la richesse spécifique aussi importante qu’en octobre mais avec une densité bien supérieure, puis de nous interroger sur la signification des variations, en juillet et en octobre.
Au mois d’août, la pelouse rase montre un groupement très particulier à Stenobothrus stigmaticus et Tetrix tenuicornis.
Les affleurements rocheux et la pelouse à Festuca lemanii ont des peuplements très voisins, ainsi que le montre la classification hiérarchique. Sur la projection F1-F3, on voit mieux leurs espèces caractéristiques, en particulier Chorthippus brunneus pour les rochers (Tetrix subulata étant mal échantillonné), les 6 autres espèces dans la pelouse (Stenobothrus lineatus, Chorthippus biguttulus, Calliptamus barbarus, Mantis religiosa, Omocestus rufipes et Platycleis albopunctata).
Par ailleurs, il y proximité entre les projections des landes mésophiles, à Fougère, et sub-sèche à Ajonc nain et Brachypode rameux. Les espèces caractéristiques sont Phaneroptera falcata, Leptophyes punctatissima et Chorthippus binotatus. Pourtant, la physionomie de ces trois groupements végétaux se reconnaît de manière évidente sur le terrain. Metrioptera roeseli, n'ayant été rencontrée qu'en un seul exemplaire, ne sera pas discutée. Enfin, la lande mésohygrophile est associée à Conocephalus discolor, Chrysochraon dispar et Chorthippus parallelus.
Au mois d'octobre, on a une bonne concordance avec le mois d’août :
-
pelouse rase à Stenobothrus stigmaticus avec adjonction d’Oedipoda caerulescens.
-
parenté entre les groupements sur rochers et pelouse dense à Festuca lemanii
-
la lande sub-sèche est caractérisée par C. binotatus et Aiolopus strepens
-
la lande à Fougère s’individualise de la lande sub-sèche par L. punctatissima et Mantis religiosa.
-
les landes mésophile et mésohygrophile se rapprochent par la présence commune de Phaneroptera falcata et Conocephalus discolor.
Au mois de juillet, on retrouve les peuplements des affleurements rocheux mais limités à Calliptamus barbarus comme espèce caractéristique. La pelouse rase comprend ici, en plus de S. stigmaticus déjà cité, Platycleis tessellata et S. lineatus. La pelouse dense à Fétuque de Leman n’abrite pas d’espèce particulière. P. albopunctata est surtout abondant dans la lande sèche, O. rufipes et M. religiosa dans la lande mésophile. Fort heureusement, L. punctatissima est fidèle à la lande mésophile. Sans surprise, M. roeseli et C. dispar sont associés à la lande mésohygrophile.
En résumé, on peut déduire de ces analyses les associations synthétiques d'Orthoptéroïdes (les astérisques indiquent les espèces pouvant changer de milieu caractéristique au cours de leur vie) :
-
Rochers : *Chorthippus brunneus
-
Pelouses rases des ouvertures : Stenobothrus stigmaticus, *Oedipoda caerulescens, Tetrix tenuicornis, Platycleis tessellata
-
Pelouse dense à Fétuque de Leman : Stenobothrus lineatus, *Calliptamus barbarus, Chorthippus biguttulus, *Omocestus rufipes, *Platycleis albopunctata, *Mantis religiosa
-
Ourlet à Brachypode et lande sub-sèche à Ajonc nain : Chorthippus binotatus, Aiolopus strepens
-
Lande mésophile : Phaneroptera falcata, Leptophyes punctatissima
-
Lande mésohygrophile : Chrysochraon dispar, Metrioptera roeseli, Conocephalus discolor, Chorthippus parallelus
Discussion et conclusion
1. Composition faunistique
La faune d’orthoptéroïdes de la lande de la Flotte et du Cluzeau comprend 24 espèces d'Orthoptéroïdes, réparties en 12 Caelifères, 11 Ensifères et 1 Mantoptère. La liste du tableau 1 de l’annexe contient, en plus des espèces citées dans les relevés, des espèces observées à l’extérieur des zones échantillonnées : la grande sauterelle verte (Tettigonia viridissima) présente dans la lande à Ajonc nain et fougère, le grillon des marais (Pteronemobius heydenii) dans les dépressions les plus humides (Bonnet et Petit, 1994), en compagnie de Chorthippus montanus.
L'absence de certaines espèces banales pour la région, comme le criquet des mouillères (Euchorthippus pulvinatus) ou le grillon italien (Oecanthus pellucens), ne trouve pas d'explication immédiate. Trois hypothèses peuvent être émises : (i) espèces ayant échappé à nos investigations ou absentes les années de prospection ; (ii) espèces dont les populations ont disparu pour une raison quelconque et non réapparues en raison de l'éloignement par rapport aux sources potentielles ; (iii) espèces phytophages défavorisées par les éléments en trace contenus dans les plantes, notamment Festuca lemanii.
- Note de bas de page 1 :
-
Son nom français “criquet des mouillères” a été bien mal choisi par G. Luquet : en Limousin, ce qriquet affectionne les zones herbeuses mésophiles à xérophiles.
Concernant Euchorthippus pulvinatus1, son absence également remarquée à Bettu (Corrèze), autre site sur serpentinite, pourrait faire penser à un problème nutritionnel. Il serait intéressant de nourrir cette espèce graminivore avec la Fétuque de Leman provenant du site pour tester si les éléments traces, en particulier le nickel, ont un impact sur sa croissance ou sa fécondité. En effet, bien que l’effet de ce métal n’ait pas été étudié chez les Orthoptères, on peut s’attendre à trouver un parallèle avec les résultats de Schmidt et al. (1991). Ces auteurs ont en effet montré qu’une augmentation de la teneur en cadmium, mercure ou plomb dans le sol diminuait significativement le nombre de pontes et la durée de vie d’Aiolopus thalassinus. Cependant, le cuivre et le cadmium peuvent être éliminés en partie avec la cuticule perdue au cours de la mue imaginale par des espèces comme Chorthippus brunneus (Hunter et al., 1987).
Le Grillon italien est discret pendant la journée et sa découverte serait facilitée par une écoute nocturne, démarche que nous avons malheureusement négligée.
Finalement, les 24 espèces d'Orthoptéroïdes de cette lande représentent 37 % des 67 espèces que compte la région Limousin (Petit et al., 2001). Trois sont considérées comme déterminantes dans les inventaires ZNIEFF du Limousin : Calliptamus barbarus, Chorthippus binotatus et Pteronemobius heydeni (DIREN, 2000). La diversité est donc moindre (plus de 30) que dans la région de Chasteaux en Corrèze (pelouses calcaires et vallées humides environnantes) (Picaud et al., 2000). Cette moins grande richesse provient de l'absence de certaines espèces de zones humides, comme Parapleurus alliaceus, Stethophyma grossum, d'espèces de pelouses écorchées comme Oedipoda germanica et Omocestus petraeus. Enfin, des espèces thermophiles comme Pezotettix giornai et Empusa pennata sont limitées par le froid.
2. Analyse des synusies orthoptériques (d'après Defaut, 1994 et 1999)
L'orthoptérofaune est constituée principalement d'espèces appartenant aux synusies euryméditerranéennes, les Oecanthea pellucentis (C. barbarus, P. tessellata, M. religiosa, A. strepens, O. caerulescens, P. tessellata) et plus particulièrement des Chorthippetea binotati (C. binotatus, C. brunneus, O. rufipes, T. tenuicornis). Le contingent d'espèces des synusies eurosibériennes (Chorthippea paralleli) est également bien représenté (S. lineatus, C. biguttulus, C. parallelus, P. albopunctata), avec en particulier les synusies de l’étage collinéen (Roeselianetea roeseli), avec Metrioptera roeselii, Chrysochraon dispar et Ruspolia nitidula. Bien que ne disposant pas d'information sur la question, P. heydenii et C. montanus seraient à placer vraisemblablement dans cet ensemble.
Les synusies des milieux arborés, les Nemobietea sylvestris, sont restreintes à peu d’espèces (N. sylvestris, L. punctatissima et P. falcata) en raison de l’absence de bois (quelques bosquets tout au plus).
Cependant, le concept de synusie est plus délicat qu'il n'y paraît. Certes, il y a une parenté avec les associations végétales et le fait de citer une liste d'espèces dans une région donnée permet souvent d'évoquer immédiatement le milieu de récolte. Quelques exemples décrits dans ce travail permettent de s'en convaincre. D’après l’analyse des résultats des AFC concernant les associations insectes-milieux, on peut penser pour certains Orthoptéroïdes à un changement dans l'écologie, avec en particulier déplacement vers les sites de ponte à la fin de la vie de l'insecte. On sait par exemple que la Mante religieuse va fixer ses oothèques sur des buissons bas, végétaux rares dans la pelouse à Fétuque mais évidemment abondants dans la lande à Fougère. En début de saison, les descendants n'ont pas eu le temps de se déplacer par rapport aux pontes de leurs parents. Au mois d’août, la Mante se déplacerait vers les pelouses denses pour regagner les buissons en fin de saison. Un même mécanisme s’appliquerait à Platycleis albopunctata, sans toutefois observer d’augmentation en début octobre dans les buissons. Oedipoda caerulescens quant à lui a besoin de milieux herbeux pendant sa vie juvénile, du moins en Limousin. Il ne devient géophile qu’une fois adulte. Les micromigrations décrites par Guéguen (1976) à propos de Stethophyma grossum, illustrent également ce phénomène. En ce qui concerne Calliptamus barbarus, on pourrait penser qu'en début de saison, il se réfugierait sur les rochers pour profiter de la chaleur. Par la suite, il occuperait préférentiellement les pelouses qui commencent à sécher. Omocestus rufipes a une position plus problématique : on peut émettre l’hypothèse que la première génération en juillet a besoin de plus d’humidité stationnelle que la deuxième génération.
Un autre article analysera les synusies de cette lande par rapport aux synusies décrites sur les coteaux calcaires (Picaud et al., 2000) et sur les landes siliceuses du Limousin (Petit et al., en préparation). Néanmoins, il semble d’ores et déjà que les peuplements de lande sub-sèche observés ici sont conformes à ce que l’on observe chez des landes comparables sur sol siliceux de la région.
En conclusion, les Orthoptères diffèrent des plantes en tant qu'outils pour la description du milieu par le fait que certaines espèces changent d'exigences écologiques au cours de leur vie. Du point de vue pratique, comme le statut d'un certain nombre d'espèces est difficile à saisir, il serait judicieux de mettre en avant les espèces fidèles aux conditions du milieu au cours de leur vie pour affiner les typologies. Du point de vue plus fondamental, nous avons encore beaucoup à apprendre de la description et de la compréhension du déterminisme de ces micromigrations.
3. Le dynamisme de végétation et l'orthoptérofaune associée
Au niveau du chaos rocheux sommital, l'abandon des pratiques agricoles donne lieu à l'apparition de la Fougère aigle et le développement de buissons de plus en plus élevés (bourdaines et chênes). On a donc :
Affleurements rocheux ⇒ pelouse écorchée ⇒ pelouse à Fétuque de Leman ⇒ pelouse à Brachypode et lande basse sèche à Ajonc nain ⇒ lande haute colonisée par les Fougères aigles ⇒ chênaie ± riche en Bourdaine.
Au niveau des peuplements, les figures 1 et 2 illustrent parfaitement la chute de la densité et de la diversité entre la lande basse à Ajonc nain et la lande plus élevée colonisée par la fougère aigle. La perte de diversité ne s’accompagne pas d’un remplacement d'espèces entre la lande à fougère et les autres landes : il n'y a pas d'espèces caractéristiques de la lande à fougère, comme nous l'avons souligné en indiquant la proximité de composition du peuplement avec la lande mésophile.
Nos observations montrent que les bois de Chêne et Bourdaine ne sont colonisés que par Ephippiger ephippiger, à une densité d'environ 1-5 pour 100 m², si l'on fait exception du grillon des bois Nemobius sylvestris.
4. Propositions de gestion
La présente étude ne préconise rien concernant les zones humides car l'essentiel de nos efforts se sont concentrés sur les peuplements des sommets. Il est certain que le dynamisme actuel de la végétation est faible car les caractéristiques chimiques et physiques du sol sont peu favorables à la croissance des plantes. Il serait cependant souhaitable de pratiquer un pâturage avec exportation de matière organique sur les parties sèches sommitales et un débroussaillage au niveau des landes mésophiles. L'intérêt serait de créer des zones de sol nu qui ne soient pas du rocher, et l'on verrait peut-être l'apparition de nouvelles espèces, comme Myrmeleotettix maculatus. Cette espèce est en effet connue pour être liée aux zones dénudées par les passages d’ovin, notamment sur la Montagne Limousine (Petit et al., en préparation). Le débroussaillage des landes mésophiles permettrait d'augmenter la biomasse des insectes et la potentialité trophique de diverses espèces de mammifères, oiseaux et reptiles.
Naturellement, ces recommandations sont d'une portée limitée car d'autres catégories d'organismes (autres ordres d'Insectes, Araignées) pourraient inciter à des mesures différentes.