Galba truncatula (O.F. Müller, 1774) : observations écologiques et données biométriques sur deux morphotypes de limnées vivant sur des sols acides Galba truncatula (O.F. Müller, 1774): ecological observations and biometric data on two morphotypes of snails living on acid soils
Des observations écologiques et des études biométriques ont été effectuées pendant trois années sur deux populations de Galba truncatula avec des coquilles présentant des tours convexes peu étagés (Châteauponsac, Chavanat) et les deux autres avec des coquilles présentant des tours en marches d’escalier (témoins). La densité des limnées tout au long de l’année était significativement plus élevée dans les populations de Châteauponsac et de Chavanat : une moyenne de 194.m-2 et de 180.m-2, respectivement, en mai ou juin, en fonction de l’altitude au lieu de 41.m-2 et de 34.m-2 dans les habitats témoins. Des résultats similaires ont également été notés pour la densité des pontes au printemps et le nombre d’oeufs par ponte. La croissance des mollusques tout au long de l’année n’a été étudiée que dans deux populations et n’a pas présenté de différence significative entre les quatre populations. Les limnées de Châteauponsac et de Chavanat ont une coquille plus étroite que celles des populations témoins, aussi bien pour la largeur du dernier tour de spire que pour celle de l’ouverture. Devant ces résultats, les auteurs proposent de classer provisoirement les deux populations de Châteauponsac et de Chavanat comme une variété de G. truncatula en attendant qu’un malacologue expérimenté en systématique précise le rang taxonomique de ces deux populations.
Ecological observations and biometric studies were carried out over three years on two populations of Galba truncatula with shells showing convex whorls without steps (Châteauponsac, Chavanat) and the other two populations with shells having characteristic step-like whorls (controls). Snail density throughout the year was significantly higher in the Châteauponsac and Chavanat populations: an average of 194.m-2 and 180.m-2, respectively, in May or June, depending on altitude instead of 41.m-2 and 34.m-2 in control habitats. Similar results were also noted for egg-mass density in spring and the number of eggs per mass. Snail growth throughout the year was only studied in two populations and showed no significant difference between the four populations. The snails of Châteauponsac and Chavanat had a narrower shell than those of the control populations, as well for the width of the last whorl as for that of the aperture. Given these results, the authors propose to temporarily classify temporarily the two populations of Châteauponsac and Chavanat as a variety of G. truncatula while waiting for studies by a malacologist experienced in systematics to specify the taxonomic rank of these two populations.
1. Introduction
La famille des Lymnaeidae comprend de nombreuses espèces de mollusques pulmonés d’eau douce avec une coquille dextre. Les membres de cette famille ont une répartition presque cosmopolite (Correa et al., 2010). Dans l’Europe de l’ouest, 13 espèces au total sont généralement admises par la plupart des auteurs (Welter-Schultes, 2012). Mais cette liste est provisoire car de nouvelles espèces de limnées comme Galba schirazensis (Küster, 1862) et Galba cubensis (L. Pfeiffer, 1839) en Espagne (Bargues et al., 2011 ; Schniebs et al., 2018) peuvent encore être découvertes lorsque les marqueurs moléculaires sont utilisés. Cet état de fait est dû à la grande diversité que les Lymnaeidae présentent dans la morphologie de leur coquille, ce qui est lié à leur plasticité éco-phénotypique importante (Samadi et al., 2000 ; Hurtrez-Boussès et al., 2005 ; Pfenninger et al., 2006). D’après Schniebs et al. (2011, 2013, 2019), cette variabilité morphologique est particulièrement nette chez les populations européennes de Radix balthica (Linnaeus, 1758), R. labiata (Rossmässler, 1835) et R. ampla (Hartmann, 1821). Ces auteurs ont émis l’hypothèse que cette variabilité serait due à une adaptation de ces espèces aux diverses conditions qui existent dans les collections d’eau selon que celle-ci est fluente ou stagnante (Schniebs et al., 2019). Il en résulte que la structure taxonomique des Lymnaeidae est plutôt compliquée et encore très controversée, avec plusieurs sous-familles, genres et sous-genres en son sein (Hubendick, 1951 ; Burch, 1982 ; Remigio et Blair, 1997 ; Jackiewicz, 1998 ; Bargues et al., 2001, 2003, 2007 ; Correa et al., 2010, 2011 ; Vinarski, 2013).
Dans le passé, de nombreux malacologues ont essayé de traduire la variation des caractères de la coquille en utilisant des catégories sous-spécifiques de rang différent, le plus souvent la « variété ». D’après Vinarski (2014), ce rang a été utilisé de manière plutôt incohérente et a été généralement appliqué à des groupes sous-spécifiques différenciés sur la base de divers traits phénotypiques (dimensions de la coquille, coloration de surface, anomalie morphologique, etc.). Cette manière de procéder a également été utilisée dans le cas de Galba truncatula (O.F. Müller, 1774). Cette limnée est la seule espèce du genre Galba à être reconnue pour l’instant par le Muséum National d’Histoire Naturelle de Paris et à être inscrite de ce fait sur la Liste de référence des Mollusques continentaux de France (Falkner et al., 2002 ; Gargominy et al., 2011). Cette décision s’appuie sur les travaux de biologie moléculaire que Bargues et al. (2001, 2003), Hurtrez-Boussès et al. (2005) et Correa et al. (2010, 2011, 2017) ont réalisés sur plusieurs populations françaises de cette espèce. Dans le passé, tous les auteurs n’étaient pas d’accord sur l’existence d’une seule espèce (G. truncatula) en France depuis les travaux de Locard (1893) ou de Germain (1903, 1930-1931). C’est ainsi que ce dernier auteur a décrit deux espèces supplémentaires de Galba et, également, deux variétés au sein de l’espèce G. truncatula (Germain, 1930-1931). Plus récemment, Adam (1960) ne liste qu’une seule espèce de Galba (G. truncatula) en Belgique, mais avec quatre variétés au sein de celle-ci. Ces variétés de G. truncatula rapportées par les auteurs précités ainsi que celles que Germain (1930-1931) a élevées au rang d’espèce ne sont probablement que des morphotypes particuliers de Limnées tronquées avec une coquille modifiée en fonction des contraintes du milieu dans lequel ces individus ont vécu.

Fig. 1. Les deux morphotypes de Galba truncatula : 1a et b, coquille assez ventrue, avec des tours se spire nettement étagés « en marches d’escalier » (individus provenant de Saint-Michel-de-Veisse, Creuse) ; 1c et d, coquille élancée avec des tours de spire peu étagés et un dernier tour peu développé (individus provenant de Châteauponsac, Haute-Vienne). Face supérieure des spécimens : 1a et 1c ; face inférieure avec l’ouverture : 1b et 1d. Hauteur de coquille des spécimens : 6,71 mm (Saint-Michel-de-Veisse), 7,92 mm (Châteauponsac).
De 1970 à 2006, notre équipe a mené plusieurs enquêtes dans 361 fermes situées dans la région du Limousin afin de détecter la présence d’habitats occupés par G. truncatula. Au total, 385,7 km2 de pâtures été prospectés dans ces exploitations et 7 709 habitats peuplés par la limnée ont été trouvés (Vareille-Morel et al., 2007 ; Rondelaud et al., 2009, 2011). Sur ces 7709 populations, 7707 ont été identifiées comme G. truncatula en se basant sur la hauteur de la coquille à l’état adulte (8 à 9 mm sur les sols cristallophylliens) et les tours de spire nettement étagés l’un par rapport à l’autre comme « des marches d’escalier » : Fig. 1a, b (Germain, 1930-1931 ; Welter-Schultes, 2012). Les deux autres populations ont présenté un autre morphotype avec un dernier tour peu développé au stade adulte et des tours de spire convexes, peu étagés : Fig. 1c, d. De plus, différentes observations faites au cours des vingt dernières années sur ces deux populations ont montré une forte densité en individus dans leurs habitats. Comme ces deux populations ont été identifiées comme G. truncatula avec les marqueurs moléculaires (Prof. S. Mas-Coma, contribution personnelle), des observations écologiques sur les caractéristiques des pontes et la dynamique de ces deux populations ont été réalisées afin de comprendre les raisons d’une telle abondance en individus. Des observations biométriques ont également été effectuées sur ces deux colonies afin de préciser les caractéristiques de leur coquille. Les résultats obtenus avec ces deux populations ont été comparés avec ceux fournis par des colonies témoins de G. truncatula, identifiées par les tours nettement étagés de leur coquille. Le but de cette étude rétrospective est seulement d’apporter des données écologiques et biométriques sur ces deux populations à morphotype particulier. Les quatre populations ont été dénommées dans le cadre de la présente note par le nom de la commune sur laquelle chaque habitat est situé.
2. Matériel et méthodes
2.1. Populations étudiées
Tableau 1 : Localisation géographique et coordonnées GPS des quatre populations de Galba truncatula dans les départements de la Creuse et de la Haute-Vienne.
|
Morphotype de Galba truncatula |
Localité |
Département |
Coordonnées GPS |
|
|
Latitude |
Longitude |
|||
|
Coquilles à tours convexes, peu étagés |
Chavanat |
Creuse |
45°57’18" N |
1°57’32" E |
|
Châteauponsac |
Haute-Vienne |
46°8’3" N |
1°17’36" E |
|
|
Coquilles à tours nettement étagés « en marches d’escalier »* |
Saint-Michel- de-Veisse |
Creuse |
45°55’47" N |
2°2’34" E |
|
Rancon |
Haute-Vienne |
46°6’56" N |
1°10’50" E |
|
* Populations témoins.
Le tableau 1 indique la localisation des quatre populations de G. truncatula et leurs coordonnées GPS. Les habitats occupés par les deux populations avec des coquilles à tours peu étagés sont situés, respectivement, sur les communes de Châteauponsac et de Chavanat. Il s’agit de fossés de route avec une altitude respective de 275 m et de 600 m. Les deux populations témoins, avec des tours de spire nettement étagés, vivent également dans des fossés de route sur les communes de Rancon et de Saint-Michel-de-Veisse. La distance entre chaque population à morphotype particulier et celle qui a servi de témoin est inférieure à 15 km. Tous les habitats sont soumis à un climat continental modulé par les vents humides qui viennent de l’Océan Atlantique. La pluviométrie annuelle moyenne fluctue de 800 à 1 000 mm selon les années, tandis que la température annuelle moyenne varie de 10° à 10,5° C (Rondelaud et al., 2009, 2011).
Le fossé de Châteauponsac longe une parcelle de terrain occupée par un immeuble commercial depuis 1995, tandis que les trois autres fossés sont situés le long de prairies pâturées. Dans chaque fossé, l’habitat occupé par les limnées est recouvert par de l’eau provenant d’une source temporaire de la mi-octobre ou de novembre jusqu’à la mi-juin (Chavanat, Saint-Michel-de-Veisse) ou de la mi-juillet (Châteauponsac, Rancon). La végétation poussant dans les deux habitats de chaque département était la même, avec une prédominance de Juncus acutiflorus Ehrhart, ex Hoffmann, 1791 dans les fossés de la Haute-Vienne et de Juncus effusus Linnaeus, 1753 dans ceux de la Creuse. Aucun escargot prédateur : Zonitoides nitidus (O.F. Müller, 1774) n’a été observé dans ces quatre fossés au cours de la période d’étude. Lorsque cet escargot est présent dans un pâturage ou une berge d’une rivière sur un sol acide, il consomme la plupart des limnées au début de l’assèchement estival des habitats (Rondelaud, 1975, 1999).
2.2. Détermination des caractéristiques des habitats
Comme les habitats des limnées sont linéaires dans les quatre fossés, leur superficie a été calculée à la mi-mars en considérant l’étendue de la nappe d’eau qui les recouvre. Le choix de cette date nous a été dicté par le fait que les limnées sont alors toutes immergées. Les limites de chaque collection d’eau sont transcrites sur une carte et la superficie globale de chaque habitat est calculée à l’aide de formules mathématiques appropriées. Comme l’étendue de chaque habitat peut varier au cours des années en fonction de la pluviométrie locale, la détermination de ce paramètre a été faite pendant trois années successives (de 2011 à 2013) à la mi-mars afin d’avoir une valeur moyenne. Dans trois cas, un mètre carré d’habitat correspondait à une longueur de fossé comprise entre 1,9 et 2,4 m, tandis que la longueur n’était que de 1,25 m dans le fossé de Chavanat.
Des prélèvements d’eau ont été effectués dans chaque fossé à la mi-mars au cours des trois années précitées. Le pH de cette eau a été mesuré à la mi-mars par la méthode électrométrique tandis que la concentration en ions calcium dissous a été déterminée par spectrophotométrie atomique.
2.3. Dénombrement des individus et des pontes
Dans les quatre habitats, des comptages réguliers de limnées ont été effectués chaque mois en 2012 afin d’étudier leur densité. Aucun décompte n’a été fait en août (Châteauponsac, Rancon) ou en juillet et août (Chavanat, Saint-Michel-de-Veisse) en raison du dessèchement estival. Dans chaque habitat, trois quadrants de 1 m² ont été sélectionnés au hasard et les individus présents dans ces zones ont été dénombrés par chasse à vue par deux personnes pendant 30 à 40 minutes. Lors du premier dénombrement, les limnées sont prélevées après le comptage par l’une des personnes et placées dans un bac extérieur afin d’éviter d’être recomptées lors du deuxième dénombrement. Ce dernier est effectué par la seconde personne afin de recenser les limnées qui auraient échappé au premier dénombrement. Les individus sont ensuite replacés sur le quadrant étudié.
Les masses d’œufs ont, de même, été décomptées tous les 15 jours pendant la période principale de pontes (avril-juin 2013) sur trois quadrants de 40 cm² sélectionnés au hasard dans chaque habitat. Après le comptage, ces pontes sont laissées en place dans chaque quadrant étudié. Aucun dénombrement n’a été effectué au cours de l’autre période de pontes (automne) car le développement de la végétation ne permettait pas un comptage précis des masses d’œufs.
Des échantillons de 50 pontes ont, enfin, été récoltés au hasard dans chaque habitat en dehors des trois quadrants précités. Ces prélèvements bimensuels ont eu lieu d’avril à juin 2013 et les pontes ont été transportées au laboratoire. Le nombre d’œufs présents dans chaque masse a été déterminé sous un stéréomicroscope (grossissement, x 16). Les pontes ont ensuite été placées individuellement dans des boîtes de Pétri de 50 mm contenant des tiges de Fontinalis sp. et de l’eau de source oxygénée en permanence (9-13 mg/L de calcium dissous dans l’eau). Après une période d’incubation allant de 4 à 10 jours, les juvéniles qui sortent de chaque ponte ont été décomptés afin de déterminer le taux d’éclosion par rapport au nombre d’œufs que celle-ci contenait.
2.4. Etudes biométriques
Des échantillons mensuels de 50 limnées adultes ont été prélevés au hasard sur une période de 12 mois (2013) dans les fossés de Chavanat et de Saint-Michel-de-Veisse. Les prélèvements n’ont été effectués que dans ces deux habitats car un herbicide avait été pulvérisé dans le fossé de Châteauponsac à la fin mai 2013, ce qui a entraîné une mortalité élevée dans cette population. La hauteur de coquille de chaque G. truncatula a été ensuite mesurée sur le terrain à l’aide d’un pied à coulisse électronique. Cette dimension est le seul paramètre à avoir été pris en compte dans cette étude car le but de celle-ci était de déterminer le nombre de générations annuelles. Les limnées ont finalement été réintroduites dans leurs habitats afin de ne pas perturber la croissance des autres individus dans ces populations.
Cinquante limnées adultes, appartenant à la génération transhivernante et en fin de vie, ont également été prélevées au hasard à la fin mai 2014 dans chacun des quatre habitats. Quatre paramètres ont été mesurés au laboratoire à l’aide d’un système d’analyse d’images Arès (Bélier, Châtillon, France).
2.5. Paramètres étudiés
Les quatre premiers ont été déterminés lors du dénombrement des limnées et de leurs pontes. Il s’agit de la densité des individus par m² d’habitat, du nombre de masses d’oeufs présentes sur 40 cm² de sédiment, du nombre d’œufs contenus dans chaque ponte et du taux d’éclosion des juvéniles à partir de ces pontes. Les valeurs individuelles notées pour chacun de ces paramètres ont été ramenées à une moyenne, avec calcul de l’écart type, en tenant compte de l’habitat étudié et de la date de prospection.
Les paramètres mesurés lors des études biométriques sont la hauteur de la coquille, la largeur maximale du dernier tour de spire, ainsi que la hauteur maximale et la largeur maximale de l’ouverture. Les valeurs des trois rapports suivants : hauteur de la coquille / largeur du dernier tour (rapport A/B), hauteur de la coquille / hauteur de l’ouverture (A/C), et largeur du dernier tour / largeur de l’ouverture (B/D), ont été également déterminées. Des moyennes avec les écarts types correspondants ont été calculées pour chacun de ces paramètres ou de ces rapports en tenant compte de l’habitat (les deux types d’étude) et de la date de prospection (l’étude sur les générations annuelles de G. truncatula).
2.6. Tests statistiques
La normalité des valeurs individuelles a été analysée pour chaque paramètre à l’aide du test de Shapiro-Wilk (Shapiro et Wilk, 1965). Comme leur distribution n’était pas normale, plusieurs tests statistiques ont été utilisés. Les tests de Scheirer-Ray-Hare et de Steel-Dwass-Critchlow-Fligner (Holländer et Wolfe, 1999) ont été utilisés pour établir la significativité des différences entre chaque population avec des coquilles à tours peu étagés et le témoin correspondant pour les paramètres suivants : densité des individus, celle des pontes et nombre d’œufs par ponte. Les différences entre les taux d’éclosion des juvéniles ont été comparées en utilisant le test Chi². Enfin, une analyse factorielle multiple a été réalisée afin de décrire et discriminer les individus selon les quatre populations étudiées et les valeurs des trois rapports. Les différentes analyses ont été réalisées à l’aide du logiciel R 3.3.0 (R Core Team, 2016).
3. Résultats
Au début du mois de mars 2012, les habitats occupés par chaque population présentant des coquilles avec des tours peu étagés et le témoin correspondant se situauent dans la même échelle de valeurs : de 16,1 à 18,5 m² en moyenne pour les habitats de la Haute-Vienne, de 23 à 24,3 m² pour ceux de la Creuse. Dans les habitats étudiés à Châteauponsac et à Rancon, le pH de l’eau courante était de 6,5 ou de 6,6, quelle que soit l’année où le pH a été mesuré, tandis que la concentration moyenne en ions calcium dissous était de 18,4 et 16,7 mg Ca++/L, respectivement. Dans ceux situés dans le département de la Creuse, les valeurs moyennes étaient légèrement inférieures : pH 6,2 et 12,7 mg Ca++/L à Chavanat, et pH 6,1 et 13,6 mg Ca++/L à Saint-Michel-de-Veisse.
3.1. Dénombrement des individus et des pontes
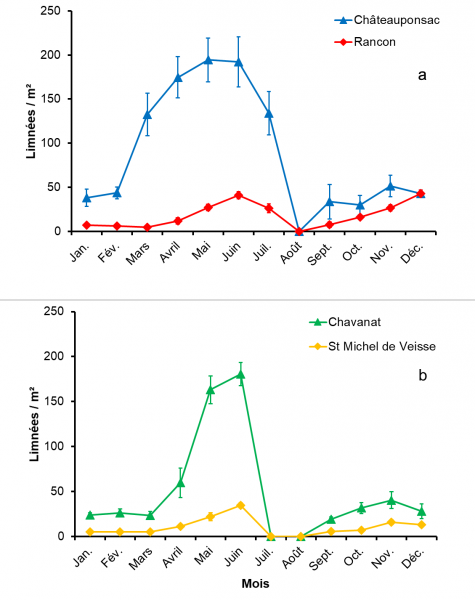
Fig. 2. Variations numériques mensuelles des limnées au cours de l’année chez les quatre populations de Galba truncatula : limnées de la Haute-Vienne (2a) et individus de la Creuse (2b). Les valeurs moyennes sont données avec leurs écarts types.
La figure 2 montre les variations numériques mensuelles des mollusques dans les quatre populations. Deux pics ont été observés dans chaque population, le premier en mai ou juin et le second en novembre, indiquant ainsi l’existence de deux générations annuelles. Dans les populations de Châteauponsac et de Chavanat, les valeurs du premier pic étaient supérieures à celles du second (respectivement 194,5.m-2 et 180,6.m-2 au lieu de 51,3.m-2 et 40,3.m-2). La même observation a également été constatée pour les populations témoins (41,6.m-2 en juin à Rancon et 34,6.m-2 à Saint-Michel-de-Veisse au lieu de 26,6.m-2 et 16,0.m-2 en novembre par ordre respectif). La localité (H = 64,71, p < 0,1 %) et la date de prospection (H = 38,76, p < 0,1 %) ont eu un effet significatif sur la distribution de ces densités. Les différences les plus significatives ont été enregistrées entre chaque population avec des coquilles à tours peu étagés et le témoin correspondant, tandis que celles existant entre les deux populations de Châteauponsac et de Chavanat, ou entre les deux témoins n’étaient pas significatives.
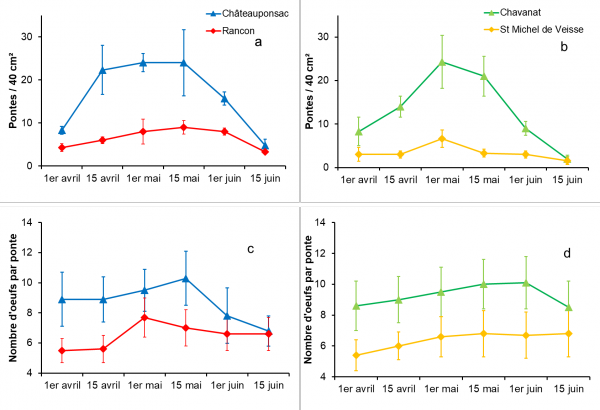
Fig. 3. Variations bimensuelles de deux paramètres chez Galba truncatula au cours des pontes printanières : la densité des masses d’oeufs sur une surface de 40 cm² (3a, b) et le nombre d’oeufs par ponte (3c, d). Les valeurs moyennes sont données avec leurs écarts types.
Les pontes déposées au printemps par les limnées transhivernantes ont montré des valeurs élevées en mai (Fig. 3a, b). Celles-ci étaient plus importantes dans les populations de Châteauponsac et de Chavanat (24,0 / 40 cm2 et 21,0 / 40 cm2, respectivement, à la mi-mai au lieu de 9,0 / 40 cm2 à Rancon et 3,3 / 40 cm2 à Saint-Michel-de-Veisse). La localité (H = 32,85, p < 0,1 %) et la date des investigations (H = 19,11, p < 1 %) ont eu, également, un effet significatif sur la distribution de ces densités. Sur chaque site, le nombre d’oeufs par masse (Fig. 3c, d) a légèrement augmenté pendant la période de pontes, avec un accroissement plus important à Châteauponsac et Chavanat que dans les sites témoins. La distribution de ces valeurs est significativement influencée par la localité (H = 490,31, p < 0,1 %) et la date des investigations (H = 66,13, p < 0,1 %).
Le taux d’éclosion des juvéniles à partir de ces œufs était de 100 % pour pontes déposées en avril et en mai. Ce taux a diminué en juin : 96,1 % au début du mois dans la population de Chavanat, par exemple, et 88,0 % à la mi-juin (données non représentées). Les différences entre les taux d’éclosion dans les quatre populations n’étaient pas significatives pour chaque date d’investigation considérée séparément.
3.2. Etudes biométriques
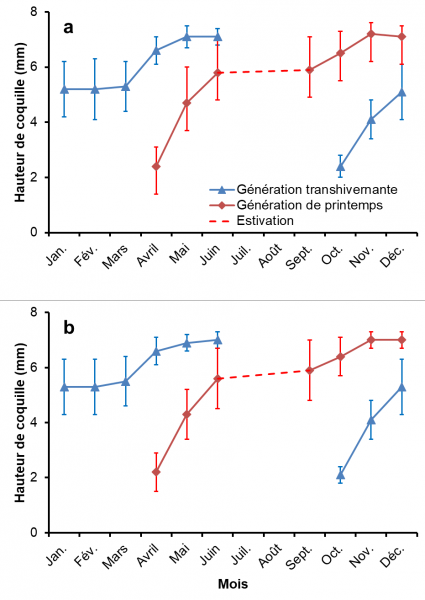
Fig. 4. Variations mensuelles de la hauteur de coquille chez deux populations de Galba truncatula au cours de l’année : Chavanat (4a) et Saint-Michel-de-Veisse (4b). Les valeurs moyennes sont données avec leurs écarts types.
La figure 4 montre la croissance des limnées dans les populations de Chavanat et de Saint-Michel-de-Veisse pour chaque mois de l’année et chaque génération considérée séparément. Dans chaque habitat, deux générations ont été notées, la première d’avril à décembre (génération de printemps) et la seconde d’octobre au mois de juin suivant (génération transhivernante). La génération de printemps a montré une croissance rapide d’avril à juin, suivie d’un plateau pendant le dessèchement estival, d’une nouvelle période de croissance de septembre à novembre et, enfin, de la disparition des derniers adultes en décembre. Après les éclosions en octobre, la génération transhivernante a montré une période de croissance rapide jusqu’en décembre, un plateau pendant les mois d’hiver (janvier-mars) et une nouvelle période de croissance au printemps jusqu’à la disparition des adultes à partir de mai. Sur les deux sites, la hauteur moyenne des coquilles des adultes en fin de vie variait de 7 à 7,2 mm. La date de prospection (H = 29,41, p < 0,1 %) a significativement influencé la distribution de ces valeurs, alors que la localité n’a eu aucun effet significatif.
Tableau 2 : Valeurs moyennes et écarts types pour quatre dimensions mesurées à la fin mai 2014 chez 200 Galba truncatula provenant de quatre populations et appartenant à la génération transhivernante. Les valeurs de trois rapports sont également indiquées. Les calculs statistiques n’ont été effectués que sur les quatre dimensions.
|
Paramètre ou rapport en mm |
Localité (département) |
|||
|
Chavanat (Creuse) |
Saint-Michel-de-Veisse (Creuse) |
Châteauponsac (Haute-Vienne) |
Rancon (Haute-Vienne) |
|
|
Hauteur de la coquille (A) |
7,91 ± 0,38 |
8,15 ± 0,49 |
8,21 ± 0,53 |
8,05 ± 0,60 |
|
Largeur du dernier tour (B) |
3,34 ± 0,29 |
4,06 ± 0,33 |
3,49 ± 0,30 |
3,92 ± 0,24 |
|
Hauteur de l’ouverture (C) |
3,59 ± 0,23 |
3,63 ± 0,26 |
3,51 ± 0,31 |
3,57 ± 0,35 |
|
Largeur de l’ouverture (D) |
2,51 ± 0,20 |
2,90 ± 0,31 |
2,52 ± 0,22 |
2.96 ± 0,26 |
|
Rapport A/B |
2,37 ± 0,12 |
2,00 ± 0,06 |
2,35 ± 0,08 |
2,04 ± 0,07 |
|
Rapport A/C |
2,20 ± 0,06 |
2,24 ± 0,05 |
2,34 ± 0,08 |
2,25 ± 0,08 |
|
Rapport B/D |
1,33 ± 0,06 |
1,40 ± 0,08 |
1,38 ± 0,06 |
1,33 ± 0,05 |
Le tableau 2 présente les valeurs moyennes et les écarts types pour les quatre dimensions que nous avons mesurées avec les limnées des quatre populations. La figure 5 montre les deux premières composantes fournies par l’analyse factorielle multiple avec les valeurs des trois rapports. Dans les quatre habitats, l’analyse permet d’expliquer 61,9 % de la variabilité totale en prenant les deux premières composantes et 81,8 % en prenant les trois composantes. Sur l’axe horizontal, la première composante est essentiellement représentée par le rapport A/B. La composante sur l’axe vertical est essentiellement représentée par les rapports A/C et B/D. La répartition des individus selon les populations est fournie sur la figure 6. Les individus des quatre habitats sont très homogènes et se mélangent peu. Ceux de Châteauponsac ont une coquille étroite avec une ouverture relativement petite par rapport à leur taille. Ceux de Chavanat ont également une coquille étroite mais l’ouverture est globalement plus large que celle des individus de Châteauponsac. Les limnées de Saint-Michel-de-Veisse et de Rancon ont une coquille globalement plus large que celle des individus de Châteauponsac et de Chavanat. Chez ces témoins, les proportions des coquilles sont assez semblables dans l’ensemble ; de même, les ouvertures de leurs coquilles sont assez proches.
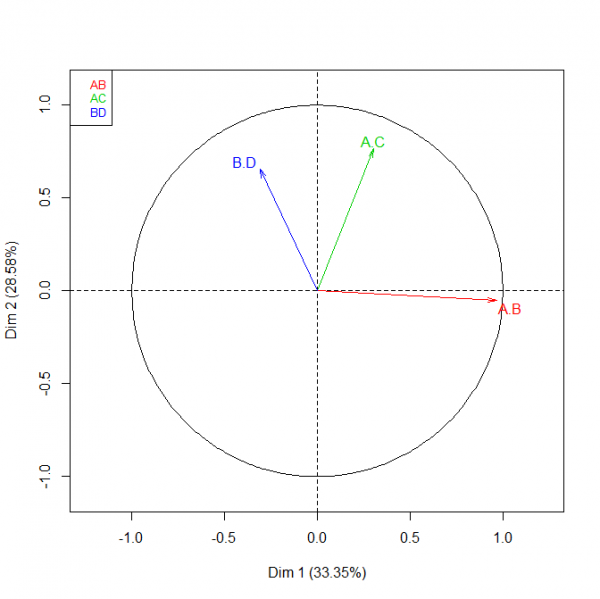
Fig. 5. Analyse factorielle multiple réalisée avec les quatre populations de Galba truncatula et les valeurs de trois rapports : contribution relative de chaque rapport à la composante. Rapports : AB, hauteur de la coquille / largeur du dernier tour de spire ; AC, hauteur de la coquille / hauteur de l’ouverture ; BD, largeur du dernier tour de spire / largeur de l’ouverture.
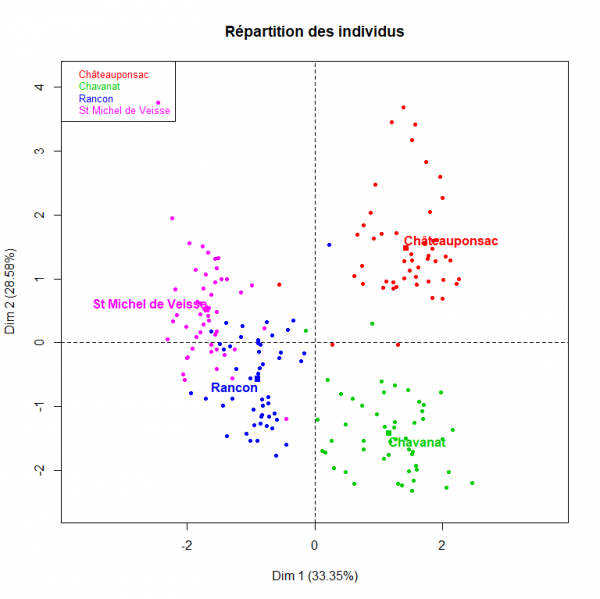
Fig. 6. Analyse factorielle multiple réalisée avec les quatre populations de Galba truncatula et les valeurs de trois rapports : répartition des individus pour chaque population.
4. Discussion
Les densités plus élevées en individus et en pontes relevées à Châteauponsac et à Chavanat indiquent la présence d’abondantes populations sur ces sites, alors que les deux habitats témoins ne sont occupés que par des populations de petite taille. Les densités observées dans les deux habitats témoins correspondent aux valeurs déjà signalées chez les G. truncatula vivant sur des sols cristallophylliens ou métamorphiques dans le centre de la France (Rondelaud et al., 2009, 2011). En revanche, les populations de Châteauponsac et de Chavanat semblent se différencier des autres populations de G. truncatula par leur densité tout au long de l’année. Les nombres d’œufs par ponte, relevés dans les quatre populations (Fig. 3c, d), sont plus difficiles à commenter car ils se situent dans l’échelle des valeurs rapportées par plusieurs auteurs pour les populations de G. truncatula sur des sols acides (Morel-Vareille, 1973 ; Rondelaud et Mage, 1992 ; Vareille-Morel et al., 1998). Comme ces nombres sont nettement inférieurs à ceux observés sur les sols sédimentaires (une moyenne de 12 à 15 œufs par ponte pour Kendall en 1953 et de 6 à 32 œufs pour Oviedo Salas en 1992), cette différence peut être expliquée par la quantité de calcium dans l’eau (généralement < 20 mg/L sur des sols acides : Guy et al., 1996) et, par conséquent, par la plus faible hauteur de coquille chez les adultes (Rondelaud et al., 2009). Cependant, le nombre d’œufs par masse est plus élevé à Châteauponsac et Chavanat que dans les populations témoins. La qualité de la nourriture présente dans les habitats des limnées (dans chaque département, les fossés ont la même végétation) et la hauteur des limnées (elle est la même au stade adulte : tableau 2) ne peut expliquer ce résultat. À notre avis, l’hypothèse la plus probable est d’admettre que les populations de Châteauponsac et de Chavanat auraient une plus grande fécondité que celle existant dans les autres populations de G. truncatula. La présence de deux générations annuelles de limées dans les deux types de populations est en accord avec les données rapportées dans la plupart des pays européens soumis à un climat tempéré (Kendall, 1953 ; Chowaniec et Drözdz, 1958 ; Heppleston, 1972 ; Morel-Vareille, 1973 ; Smith, 1981, par exemple).
Les résultats de l’analyse factorielle multiple démontrent nettement que les coquilles étudiées chez les limnées de Châteauponsac et de Chavanat se démarquent nettement des coquilles étudiées dans les deux populations témoins car elles sont plus étroites comme en témoigne la largeur du dernier tour de spire et celle de l’ouverture (Tableau 2). Ce type de morphotype est rare sur les sols acides du Limousin (2 populations sur un total de 7709 identifiées au cours de notre enquête : Rondelaud et al., 2009, 2011). La présence d’autres populations présentant ce morphotype sera probablement rare dans le département de la Haute-Vienne (c’est celui que notre équipe a le plus étudié au cours de ces 50 dernières années). Par contre, il n’en est pas de même pour la Creuse car la plupart des 56 fermes prospectées au cours de notre enquête se situaient dans le nord ou l’ouest du département et seules quelques exploitations ont fait l’objet d’investigations malacologiques dans les régions d’Aubusson et de Bourganeuf (Vignoles et al., 2017). Il est donc possible que d’autres populations avec des coquilles à tours peu étagés soient identifiées dans ce département dans les années à venir, en particulier dans la zone est. La rareté de ces populations soulève, de plus, un problème car on ne peut déterminer si les fortes densités relevées dans les deux habitats doivent être rapportées à ce morphotype ou s’il s’agit d’un processus local indépendant du morphotype, même si cette abondance d’individus a été constatée par notre équipe dans les deux habitats depuis les années 2000.
5. Conclusion
Les deux populations de Châteauponsac et de Chavanat, présentant des coquilles avec des tours peu étagés, se différencient nettement des autres populations de G. truncatula par la largeur plus faible du dernier tour de spire et par leurs effectifs élevés. Comme ces deux populations ont été identifiées comme G. truncatula à l’aide des marqueurs moléculaires, ce morphotype particulier pourrait être considéré provisoirement comme une variété phénotypique au sein de l’espèce. La morphologie de ces coquilles nous permet d’émettre l’hypothèse que les individus de ces deux populations pourraient appartenir à la variété montana (Bourguignat, 1888), variété que Germain (1930/1931) considèrait comme la forme montagnarde de G. truncatula en France. La détermination du rang taxonomique pour ces deux populations nécessite l’assistance d’un malacogue spécialisé en systématique après une étude de la variabilité génétique intra-spécifique chez G. truncatula.