Biodiversité des orthoptères dans une petite tourbière des monts d’Ambazac (Haute-Vienne) en relation avec la dynamique végétale Biodiversity of orthoptera in a small peat bog in monts d’Ambazac (Haute-Vienne) in relation with vegetal dynamics
Les auteurs analysent la dynamique végétale et les peuplements d’Orthoptères de la tourbière de la Crouzille (Monts d’Ambazac, Haute-Vienne). Les épaisseurs de tourbe sont mises en relation avec la végétation actuelle. Le fond tourbeux abrite une faune orthoptérique originale, les espèces les plus abondantes étant Chorthippus montanus et Conocephalus discolor. Au cours de l‘été, le milieu qui maintient le plus la diversité des Orthoptères est la cariçaie à Carex rostrata. Les stades plus évolués de la dynamique végétale subissent une forte érosion en terme de diversité et de densité d’insectes. Cette cariçaie est la formation dont la conservation est prioritaire.
The authors analyze vegetal dynamics and Orthoptera communities of Crouzille’s peat bog (Monts d’Ambazac, Haute-Vienne, France). The depths of dig peat were related to present vegetation. The peaty shallow is characterized by an original orthopteran fauna, the most abundant species being Chorthippus montanus and Conocephalus discolor. During summer, Carex rostrata formation is the environment keeping the highest diversity of Orthoptera. Later stages in the vegetal dynamics undergo dramatic erosion in term of insect diversity and density. Conservation measures should concern the Carex rostrata formation in priority.
Introduction
Les zones humides sont menacées constamment par les activités anthropiques alors qu’elles représentent un intérêt écologique non négligeable. Elles jouent notamment un rôle tampon dans l’écoulement des eaux. Les zones de tourbières sont parmi les plus rares et les plus intéressantes des zones humides car leur formation nécessite des conditions très particulières et le milieu qui est ainsi généré a lui-même des conditions très spécifiques de température, d’humidité, de substratum. La diversité des biotopes leur confère une faune assez remarquable tant par leur diversité que par que par la présence d’espèces à grande valeur patrimoniale comme le lézard Zootoca vivipara ou les libellules du genre Somatochlora.
Les tourbières du Limousin occupent une surface d’environ 7500 hectares, soit environ 0,45 % du territoire. Plus de 550 sites tourbeux ont été recensés, répartis sur 145 communes. Les mieux connues dans la région Limousin, des ZNIEFF de type 1, sont incontestablement la tourbière du ruisseau des Dauges (réserve naturelle nationale à Saint- Léger-La-Montagne, 87) et la tourbière du Longeyroux (arrêté de protection de biotope à Meymac, 19). Les surfaces occupées par les alvéoles sont importantes : environ 50 hectares, dont 30 de milieux tourbeux pour les Dauges et 255 hectares pour le Longeyroux.
La tourbière de la Crouzille (commune de St-Sylvestre) est une tourbière de petite taille de la bordure ouest des Monts d’Ambazac, située à moins de 10 Km de la tourbière des Dauges. Il s’agit d’une tourbière ombrogène, c’est-à-dire dont l’alimentation se fait principalement par les eaux de pluie et par le ruisseau des Sagnes.
Nous nous proposons d’étudier les peuplements d’Orthoptères en relation avec les groupements végétaux. Cette mise en parallèle des peuplements entomologiques/végétaux permettra de prévoir l’évolution temporelle de la biodiversité des peuplements d’insectes, puisque la dynamique végétale des tourbières est bien établie (BOTINEA U et al., 1998). Par ailleurs, la mise en évidence des stades les plus remarquables nous fournira des éléments pour préconiser des méthodes de conservation.
Matériel et méthodes
1- Présentation du site
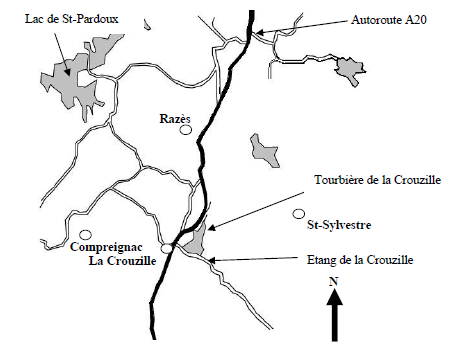
L’étang de la Crouzille est situé en bordure ouest des Monts d’Ambazac, sur la commune de Saint-Sylvestre, à une vingtaine de kilomètres de Limoges (figure 1). La zone précise de notre étude est juste en amont de l’étang de la Crouzille, au nord de ce dernier. Elle est traversée par le ruisseau des Sagnes qui constitue l’alimentation principale de la retenue.
Figure - 1. Localisation du site d’étude
L’altitude du site est d’environ 430 mètres (au plus bas). Il est constitué d’une zone de dépression avec un fond quasiment plat et des pentes la bordant. La zone tourbeuse se situe dans la dépression. Les sommets l’entourant culminent à environ 500 mètres.
La superficie du site est d’une douzaine d’hectares toutes formations végétales comprises, la zone humide n’occupant qu’une portion. Il fait partie de la ZNIEFF de type 1 de l’étang de La Crouzille, elle-même comprise dans la ZNIEF de type 2 des Monts d’Ambazac.
L’étymologie même du site montre que nous sommes en présence de zones marécageuses, puisque Sagnes viendrait du gaulois sagna ou du bas-latin sania, signifiant marais (FALC’HUN, 1981).
L’alimentation se fait principalement par les eaux de pluie et par le ruisseau. Il faut en fait distinguer deux bassins versants qui alimentent la zone : le grand bassin versant du ruisseau des Sagnes et un petit bassin au nord-ouest (fig. 2). Leur surface est respectivement de 235 ha et 45 ha. Les débits moyens correspondant à chaque bassin sont de 60 L/s pour le grand bassin et de 8 L/s pour le petit.
2- Epaisseur de tourbe
La mesure de l’épaisseur de tourbe à différents points du site permet d’évaluer sa distribution dans la tourbière. Nous avons utilisé pour cela une pige graduée de 3 mètres de longueur totale. A chaque relevé nous l’enfonçons jusqu’à atteindre une couche dure où il nous est impossible de continuer. La hauteur de la couche meuble est ainsi mesurée : sédiments fins à proximité d’un ruisseau ou tourbe.
3- Groupements végétaux
Nous avons fondé notre méthode sur la méthodologie des inventaires phytosociologiques de Braun-Blanquet. Cela consiste à dresser la liste des plantes présentes dans une surface au peuplement homogène. Nous affectons à chacune le coefficient de abondance-dominance de la phytosociologie, selon 7 notations différentes :
-
5 : 75 %-100 % de recouvrement
-
4 : 50 %-75 %
-
3 : 25 %-50 %
-
2 : 10 %-25 %
-
1 : 1 %-10 %
-
+ : <1 %
-
i : individu isolé
Avec cette méthode, nous pouvons dégager les espèces caractéristiques de chaque milieu et ainsi les définir au mieux pour les comparer avec le plus de rigueur possible.
Pour la réalisation de cet inventaire, huit sorties sur le terrain ont été effectuées :
10/12/02, 06/01/03, 16/01/03, 14/02/03, 26/02/03, 28/02/03, 10/03/03 et 23/05/03.
4- Relevés d’Orthoptères
Les sorties consacrées aux Orthoptères ont eu lieu les 25/07/03 et 02/09/03. Le temps lors de ses sorties était ensoleillé donc propice aux observations. La première a eu lieu en début de saison pour les Orthoptères afin de quantifier la diversité et la densité maximales. La seconde a eu lieu en fin de saison pour voir l’évolution du peuplement à l’issue de la période de sécheresse de l’été 2003 et pour trouver éventuellement des espèces plus tardives.
Les recensements d’Orthoptères ont été effectués sur des surfaces constantes de 9 m² à végétation uniforme. Il faut répéter 5 fois ce protocole par formation végétale et cumuler les résultats pour pouvoir avoir une bonne idée des peuplements. A chaque recensement, tous les individus s’y trouvant la zone de 9 m² sont systématiquement identifiés puis relâchés à distance. Nous obtenons ainsi une liste complète des espèces avec leur effectif.
Afin de minimiser les risques de contamination entre deux relevés, les périmètres de recherches sont éloignés d’au moins 10 mètres. Le phénomène de bordure est quant à lui évité par une prise de distance de quelques mètres par rapport aux autres formations végétales.
L’analyse factorielle des correspondances a été effectuée grâce au logiciel PAST vers 1.34 (HAMMER et HARPER, 2001).
Résultats
1- Sondages de tourbe
Les résultats des sondages sont reportés sur la figure 2. Tout d’abord, nous observons près des cours d’eau des profondeurs comprises entre 120 et 150 cm. Ces épaisseurs assez importantes correspondent ici à des sédiments fins dus au ruisseau et non à de la tourbe. Cependant, nous ne pouvons pas exclure qu’il y ait des vestiges tourbeux sous ses alluvions.
Figure 2 - Topographie de la tourbière de la Crouzille. Equidistance des courbes de niveaux : 10 m. Ellipses gris clair : profondeur de tourbe entre 150 cm et 180 cm ; ellipse gris foncé : entre 200 et 250 cm.
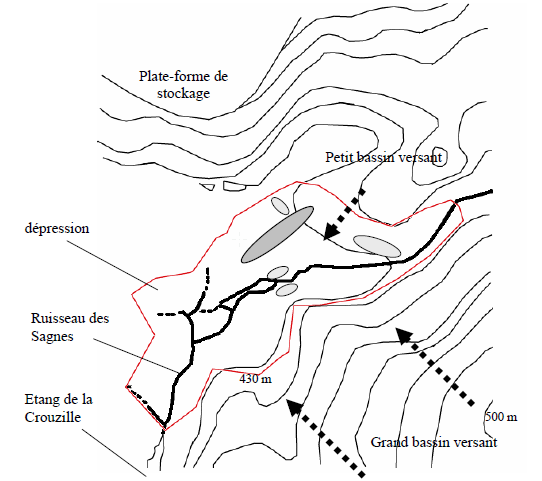
Les autres mesures représentent réellement des épaisseurs de tourbe. Elles sont très variables et se situent entre 70 cm et 250 cm. La partie où la tourbe est la plus profonde forme un axe orienté SSO-NNE. Cela pourrait s’expliquer par plusieurs phases de creusement au cours du quaternaire comme cela a été montré au Longéroux ; le long de cet axe, les sphaignes se sont installées et ont produit de la tourbe. Petit à petit, le trou s’est comblé puis bombé tout en gardant ses caractéristiques humides et a « contaminé » ses abords. La formation de la tourbe dans ces derniers est donc plus récente et comme cela se joue sur des centaines d’années, la tourbe y est moins épaisse. La tourbière « originelle » du site dans le contexte actuel doit probablement se trouver là où les profondeurs sont maximales. Ceci ne peut rester qu’une hypothèse car sur un même site, les épaisseurs de tourbe peuvent être très différentes pour un même âge (MANNEVILLE et al., 1999). Dans ce cas, ce sont les variations sensibles du milieu ainsi que les différentes phases d’activité qui expliquent les différences d’épaisseurs.
2- Flore et végétation
Chaque relevé contient les espèces rencontrées avec leur plus fort coefficient d’abondance constaté selon les saisons. 99 relevés ont été effectués au total.
Les prospections nous ont permis de nous rendre compte d’un premier fait : les groupements végétaux sont relativement nombreux et leur succession est parfois assez floue. Ils ne sont pas homogènes et peuvent varier dans les proportions en leurs différentes espèces. Les noms des groupements sont conformes à ceux utilisés dans l’Atlas de la flore vasculaire en Limousin (BRUGEL et al., 2001).
Nous avons recensé 102 espèces végétales, en excluant les Bryophytes excepté Polytrichum commune. Il sous-estime notamment les Poacées que nous n’avons pas toutes déterminées.
Pour comparaison, la Réserve Naturelle de la tourbière des Dauges comprend 207 plantes vasculaires pour une superficie de 200 ha environ (BOTINEAU et al., 1998 ; CRENL, 2000). La différence significative entre ces deux tourbières est due sans doute à l’écart de superficie, à une plus grande diversité de milieux (en particulier les landes sèches) et à la détermination systématique, sur plusieurs années, de toutes les espèces présentes à la tourbière des Dauges.
Il faut remarquer ici qu’aucune espèce protégée au niveau national n’a été rencontrée. Toutefois, nous avons 8 espèces à grand intérêt patrimonial pour le Limousin car en limite d’aire de répartition (DIREN, 1999) : Carex nigra, Carex rostrata, Ceratocapnos claviculata, Menyanthes trifoliata, Narthecium ossifragum, Potentilla palustris, Viola palustris, Wahlenbergia hederacea. Hormis Ceratocapnos claviculata, toutes sont des espèces strictement de milieu humide et plus précisément de milieu tourbeux.
2.1. Formations ligneuses
Les saulaies constituent la formation dominante, mais cette dernière connaît quelques déclinaisons. En effet, elle peut être franchement marécageuse, le long des cours d’eau ou relativement plus sèche avec une prairie au pied. Les saulaies humides sont considérées comme des bois hygrophiles appartenant à l’alliance phytosociologique du Salicion cinereae, classe des Alnetea glutinosea. Il faut noter ici que les formations à bourdaine et à bouleau font partie de ces bois hygrophiles. Nous y retrouvons notamment Carex paniculata, Agrostis canina, Galium palustre, Dryopteris carthusiana et diverses sphaignes. Ce sont des formations tourbeuses relativement peu représentées sur le site, limitées à la frange est. Ces bois tourbeux à sphaignes sont considérés dans la Directive Habitats comme prioritaire en tant que tourbière boisée, code Natura 2000 91DO (Ministère de l’agriculture et de la pêche et al., 2002). Dans certains cas, Carex paniculata constitue des groupements sub- monospécifiques de plus de 100 m². La formation à saule la plus importante est représentée par les saulaies marécageuses à Caltha palustris, Rubus fruticosus, Dryopteris carthusiana, Galium palustre, Solanum dulcamara, qui appartiennent à l’alliance phytosociologique de l’Alnion incanae. Nous pouvons y retrouver de la bourdaine et du tremble. C’est ce type de formation que l’on trouve le long des cours d’eau sur substrat vaseux et mouillé.
Des groupements végétaux de bois sont aussi présents en périphérie de la zone humide avec notamment des bois de chênes, de châtaigniers ou bien des hêtraies. Les deux premiers appartiennent à l’ordre des Quercetalia robori-petraeae. On y trouve les accompagnant, Ilex aquifolium, Teucrium scorodonia, Deschampsia flexuosa, Pteridium aquilinum, Lonicera periclymenum, Hedera helix entre autres. Ces bois acidiphiles typiques se classent dans l’alliance du Quercion robori petraeae. Bien que fréquent en Limousin, ce type forestier est mal défini quant à l’association ou les associations le constituant. Les hêtraies appartiennent ici à l’ordre des Fagetalia sylvaticae, alliance du Luzulo luzuloidis-Fagion sylvaticae, sous- alliance de l’Ilici aquifolii-Fagenion sylvaticae. Ces bois représentent le stade climacique sur les moyens plateaux limousins, bien qu’ils aient été souvent remplacés, par action anthropique, par des chênaies de substitution sub- ou para- climaciques. Ces trois derniers types de boisement que nous avons sur le site représentent des stades ultimes de l’évolution végétale des pentes sur les moyens plateaux. Ils constituent donc des formations végétales qui n’évoluent plus ou très peu d’un point de vue botanique si elles sont laissées en l’état.
2.2. Formations herbacées
Hormis les parties boisées et les cours d’eau, la zone humide abrite trois formations végétales différentes d’un point de vue phytosociologique. La première est la jonçaie- moliniaie et la moliniaie tourbeuse qui appartiennent à l’association du Caro verticillati – Molinietum caeruleae. Ces prairies tourbeuses se classent plus généralement dans l’ordre des Molinetelia caeruleae, alliance du Juncion acutiflori. Ce type de formation est représenté sur le site principalement à l’ouest, entre le talus et les boisements. Il peut être la résultante d’une dégradation du milieu tourbeux (MANNEVILLE et al., 1999 ; Ministère de l’agriculture et de la pêche et al., 2002). En effet, un assèchement partiel peut conduire à ce type de groupement.
La deuxième est la mégaphorbiaie à Juncus acutiflorus et Angelica sylvestris, accompagnée par la molinie (Molinia caerulea), Cirsium palustre, Lysimachia vulgaris, Galium palustre et Galium uliginosum. L’association de ces mégaphorbiaies correspond au Junco acutiflori – Angelicetum sylvestris. Cette formation, assez réduite, est considérée sur la carte établie par LOLIVE (2003) comme une moliniaie, en raison de zones de transition entre ces deux premières formations. Nous les distinguons dans le texte pour une plus grande rigueur.
La troisième est la cariçaie tourbeuse à Carex rostrata. Cette laîche constitue l’essentiel de la végétation dont le tapis continu de sphaignes s’est épaissi et s’accroît par le haut. L’abondance des autres espèces reste modeste. C’est un groupement voisin des tourbières bombées actives de la classe des Oxycocco palustris – Sphagnetea magellanicae et de l’ordre des Erico tetralicis – Sphagnetalia papillosii. Ce groupement est souvent rattaché aux groupements des bords d’étang, alliance du Magnocaricion elatae, ordre des Magnocaricetalia elatae, classe des Phragmito australis – Magnocaricetea elatae. Ce groupement occupe principalement l’espace entre les deux groupements à saules et représente une superficie non négligeable.
Cette formation correspond au groupement de tourbières hautes actives (code 51.1) de la Directive Habitat, dans laquelle elle est considérée comme habitat prioritaire. Le classement de ce groupement parmi ceux qui sont les plus importants à sauvegarder confère au site un intérêt écologique très important. Cependant, si cette formation est non négligeable sur le site, elle reste d’une faible superficie (moins d’un hectare), ce qui en limite son intérêt.
2.3. Evolution temporelle des formations végétales
Fort de nos constatations et de nos inventaires (Annexe), nous avons essayé de dégager les grands traits de l’évolution du milieu dans les dernières décennies pour mieux envisager ses transformations futures. Pour cela nous avons tenu compte des photographies aériennes prises par la COGEMA en 1970. Celles-ci nous permettent d’apprécier l’évolution du site sur une trentaine d’années et d’avancer des hypothèses quant à la dynamique des formations végétales. De plus nous avons trouvé des indices de cette dynamique sur le terrain et des pistes dans la bibliographie.
Nous avons donc pu nous rendre compte que les formations à saules sont anciennes même si la taille des arbres reste modeste. Cependant, à la lumière des photographies aériennes, nous constatons une avancée de ces saules sur le reste des formations et notamment sur les cariçaies à Carex rostrata et les prairies tourbeuses (moliniaie). Les saulaies sont probablement apparues le long des cours d’eau car ce sont des zones plus riches en éléments nutritifs et en matériaux fins et progressent de part et d’autres de ceux-là. Cette évolution vers un boisement de la zone est normale et irrémédiable mais se fait de manière très lente compte tenu des conditions difficiles qui y règnent. Dans ces saulaies, l’épaisseur des sédiments est d’environ 130 cm. Cette épaisseur relativement importante est le fait d’une accumulation de particules fines. Comme il a été dit précédemment, il pourrait s’agir en profondeur d’une ancienne accumulation de tourbe recouverte ensuite par les alluvions du ruisseau.
Les saulaies semblent aussi gagner sur les cariçaies à Carex paniculata qui ont un fonctionnement particulier. En effet, la présence de cette plante signale une fluctuation du niveau de l’eau (MANNEVILLE et al., 1999). Le substrat étant recouvert d’une tranche d’eau de plusieurs centimètres à certaines périodes et simplement humide durant les périodes plus sèches. Par effet de lisère, les saules peuvent coloniser ce milieu mais de manière très lente car les touradons ont des dimensions très importantes, jusqu’à 1,50 mètre, et sont proches les uns des autres, ce qui empêche les autres espèces de se développer. Ce sont donc des formations quasi monospécifiques. L’épaisseur du substrat est la même que dans les saulaies et a probablement la même origine. Des différences doivent tout de même exister, pour expliquer la différence de végétation. Nous constatons même des Carex paniculata dans les saulaies marécageuses ce qui est très fréquent en Limousin (BRUGEL et al., 2001). Or, nous avons aussi des zones où les touradons de Carex paniculata sont en train de mourir ou sont déjà morts sous des saulaies. L’explication de ce fait assez singulier réside sans aucun doute dans le surcreusement du lit du ruisseau à l’ouest de la levée de terre et ce jusqu’à l’étang. Ce surcreusement a provoqué un certain assèchement des zones les plus proches, ne leur permettant plus d’être inondées une partie de l’année, ce qui a condamné les Carex paniculata.
Dans les zones humides, les saulaies ne sont pas amenées à évoluer beaucoup. Cependant en périphérie, du fait d’une surélévation légère du substrat entraînant un certain assèchement, nous notons l’apparition d’une zone de transition à Carex laevigata. Cette zone est souvent assez étroite, de l’ordre du mètre. La surélévation s’accentuant, nous avons l’apparition des formations boisées climaciques ou subclimaciques. Nous avons des hêtraies sur les pentes à exposition fraîche et des bois mixtes (chênaies, châtaigneraies) quand les conditions sont plus douces. La présence de ces deux formations n’est peut-être pas uniquement le fait de l’exposition, mais également de l’action de l’homme. Les bois mixtes remplacent souvent les hêtraies après que celles-ci aient été déboisées.
En ce qui concerne les zones que nous avons précédemment qualifiées de tourbière active, leur évolution est assez facile à prévoir. Les zones à Carex rostrata, comme nous l’avons déjà vu, ont tendance à se boiser mais de manière très lente à cause des conditions rigoureuses du milieu et du recouvrement très important du substrat par les sphaignes empêchant ainsi la germination des ligneux (MANNEVILLE et al., 1999). Ces milieux tourbeux ont des épaisseurs de tourbe comprise entre 120 cm et 180 cm. Ces variations sont normales car la fabrication de la tourbe est plus ou moins importante selon les conditions du milieu qui sont relativement variables même dans cette zone d’apparence homogène. La formation de tourbe dans ces zones est a priori plus récente que dans les zones de tourbière bombée évoluée. Cependant, grâce à la quantité importante de sphaigne présente, la turbification est encore importante entraînant un épaississement continuel de la tourbe. Cet état de fait durera autant que les sphaignes perdureront sur cette zone et que les ligneux resteront dans de très faibles proportions.
Cependant l’évolution vers une tourbière bombée évoluée est obligatoire et se fait par une augmentation des ligneux et sous-ligneux. Le premier à s’installer est la bruyère à quatre angles (Erica tetralix) et quelques pieds poussent déjà dans ces cariçaies. Puis nous aurons l’installation de ligneux plus grands comme les saules et les bouleaux, mais aussi la bourdaine. Ce stade d’évolution que nous pouvons presque qualifier d’ultime, est présent sur notre site avec la petite boulaie centrale et une partie de la boulaie et du bois à bourdaine de la frange est. C’est dans ces formations de tourbière bombée évoluée que nous observons les plus grandes épaisseurs de tourbes avec plus de 2 mètres (figure 2). Ces épaisseurs importantes, mises en correspondance avec la végétation, suggèrent que ce sont les zones turfigènes les plus anciennes du site. Cependant, l’épaisseur de tourbe n’est pas toujours proportionnelle à l’âge de la tourbière (MANNEVILLE et al., 1999). Nous ne pouvons donc pas expliquer ici ces grandes variations d’épaisseurs du substrat. Dans ces formations, même si nous avons encore des sphaignes en quantité importante, il est difficile de quantifier la production de tourbe.
Les prairies tourbeuses constituées par des moliniaies et des jonçaies-moliniaies subiront également une évolution vers un boisement, d’autant plus rapide qu’elles ne sont pas exploitées. Dans ces formations, l’épaisseur de tourbe est moyenne (130 cm) et n’évolue probablement pas très vite du fait de la faible quantité de sphaignes. Ces formations peuvent être le résultat d’une dégradation d’un milieu franchement tourbeux par assèchement ou par exploitation à visées agricoles, comme le pâturage ou le fauchage (Ministère de l’agriculture et de la pêche et al., 2002). Si de telles pratiques ont eu lieu sur le site, aucune trace n’en subsiste ; nous ne pouvons donc qu’en rester au stade des hypothèses.
Les relations supposées entre les différentes formations végétales sont indiquées de manière synthétique dans la figure 3.
La dynamique végétale du site est donc relativement complexe du fait de la multitude des formations présentes. Cependant, l’évolution naturelle vers un boisement a été constatée en comparant la situation d’aujourd’hui et celle d’il y a une trentaine d’années. Cette évolution est normale et se fait sur des dizaines voire des centaines d’années compte tenu des conditions très difficiles de ce milieu (MANNEVILLE et al., 1999).
Figure 3. - Essai interprétatif de la dynamique végétale
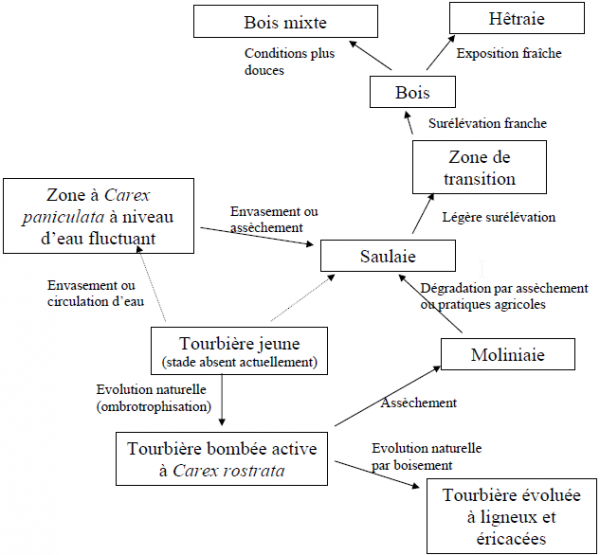
Légendes : trait continu = déterminisme probable ; tirets = déterminisme inconnu
3- Faune Orthoptérique
Les densités des espèces rapportées à 100 m² sont reportées dans les tableaux I et II. En juillet, la tourbière bombée active (zone à Carex rostrata) montre le peuplement en insectes le plus dense avec plus de 200 individus pour 100 m². Il s’agit également de la zone la plus diversifiée parmi les formations de milieux humides. C’est là où nous trouvons la faune la plus spécialisée, avec en particulier Chorthippus montanus. Ce criquet a tendance à se raréfier dans les zones humides du nord de la France et remplace C. parallelus, une espèce très commune, dans les parties les plus humides. Par rapport au Dauges (BONNET et PETIT, 1998), plusieurs espèces n’ont pas été trouvées (Metrioptera brachyptera, Pteronemobius heydenii) mais la poursuite de l’inventaire dans le futur permettra de compléter la liste actuelle.
|
Juillet |
Code |
Cr |
Mo |
Ju |
Cp |
Cre |
SJ |
Zp |
somme |
|
Aiolopus thalassinus |
Aiotha1 |
0 |
0 |
0 |
0 |
0 |
0 |
13,3 |
13,3 |
|
Chorthippus albomarginatus |
Choalb1 |
0 |
0 |
0 |
0 |
0 |
0 |
13,3 |
13,3 |
|
Chorthippus montanus |
Chomon1 |
64,4 |
13,3 |
20,4 |
8,9 |
61,1 |
0 |
0 |
168,1 |
|
Chorthippus parallelus |
Chopar1 |
2,2 |
31,1 |
16,7 |
0 |
0 |
0 |
46,7 |
96,7 |
|
Chrysochraon dispar |
Chrydis1 |
6,7 |
20 |
20, |
0 |
0 |
2,2 |
0 |
49,3 |
|
Conocephalus discolor |
Conodis1 |
111.1 |
46,7 |
9, |
2,2 |
16,7 |
4,4 |
2,2 |
192,6 |
|
Leptophyes punctatissima |
Leptpun1 |
0 |
2.2 |
1,9 |
0 |
0 |
0 |
0 |
4,1 |
|
Metrioptera roeselii |
Metroes1 |
33,3 |
13,3 |
0 |
0 |
0 |
0 |
2.2 |
48,8 |
|
Oedipoda caerulescens |
Oedcae1 |
0 |
0 |
0 |
0 |
0 |
0 |
4,4 |
4,4 |
|
Omocestus rufipes |
Omoruf1 |
0 |
0 |
0 |
0 |
2,8 |
0 |
6,7 |
9,5 |
|
Omocestus viridulus |
Omovir1 |
0 |
0 |
0 |
0 |
0 |
0 |
6,7 |
6,7 |
|
Phaneroptera falcata |
Phafal1 |
2,2 |
0 |
0 |
0 |
0 |
0 |
0 |
2,2 |
|
Pholidoptera griseoaptera |
Phogri1 |
2,2 |
0 |
0 |
2,2 |
2,8 |
6,7 |
0 |
13,9 |
|
Stethophyma grossum |
Stegro1 |
0 |
0 |
3,7 |
0 |
0 |
0 |
2,2 |
5,9 |
|
Densités pour 100 m² |
222,1 |
126,6 |
72,4 |
13,3 |
83,4 |
13,3 |
97,7 |
Tableau I. - Densité des Orthoptères dans les différentes formations végétales au mois de juillet. Abréviations : Cr : Carex rostrata, Mo : Molinia caerulea, Ju : Juncus sp., Zp : Zone perturbée, Cp : Carex paniculata, Cre : C. rostrata évolué, SJ : Sparganium + Juncus.
|
Septembre |
Code |
Cr |
Mo |
Ju |
Cp |
Cre |
SJ |
Zp |
somme |
|
Aiolopus thalassinus |
Aiotha2 |
0 |
0 |
0 |
0 |
0 |
0 |
2,22 |
2,22 |
|
Chorthippus biguttulus |
Chobig2 |
0 |
0 |
0 |
0 |
0 |
0 |
6,67 |
6,67 |
|
Chorthippus brunneus |
Chobru2 |
0 |
0 |
0 |
0 |
0 |
0 |
26,67 |
26,67 |
|
Chorthippus montanus |
Chomon2 |
37,78 |
1,85 |
0 |
4 |
31,11 |
0 |
0 |
74,74 |
|
Chorthippus parallelus |
Chopar2 |
0 |
0 |
0 |
0 |
0 |
0 |
2,22 |
2,22 |
|
Chrysochraon dispar |
Chrydis2 |
0 |
0 |
0 |
4 |
0 |
0 |
0 |
4 |
|
Conocephalus discolor |
Conodis2 |
6,67 |
5,55 |
6,67 |
0 |
6,67 |
2,22 |
2,22 |
30 |
|
Omocestus rufipes |
Omoruf2 |
0 |
0 |
0 |
0 |
0 |
0 |
2,22 |
2,22 |
|
Pholidoptera griseoaptera |
Phogri2 |
0 |
0 |
0 |
1,33 |
0 |
0 |
0 |
1,33 |
|
densités pour 100 m² |
44,45 |
7,40 |
6,67 |
9,33 |
37,78 |
2,22 |
42,22 |
Tableau II. - Densité des Orthoptères dans les différentes formations végétales au mois de septembre. Abréviations : voir tableau I.
Lorsque la zone à Carex rostrata évolue, la densité chute brutalement (moins de 100 individus par m²) et nous notons la disparition de trois espèces (C. parallelus, M. roeseli et P. falcata). Cependant C. montanus se maintient. Le boisement par la bourdaine ou le bouleau entraîne une disparition complète de ces insectes.
La zone perturbée (remblais en contre bas de la plateforme de stockage) présente neuf espèces dont une bonne partie a une large répartition en France, comme C. albomarginatus, Omocestus rufipes et Oedipoda caerulescens. Cependant, il est intéressant de constater la présence d’un criquet peu commun en Limousin Aiolopus thalassinus, considéré comme une espèce déterminante (DIREN, 1999).
Les densités ont fortement en septembre chuté dans tous les milieux, ce qui est normal car d’une manière générale, les densités diminuent de moitié entre le mois de juillet et le mois d’août et de même entre août et septembre. Cependant, l’effet de la canicule estivale s’est fait significativement sentir sur les moliniaies et les jonçaies qui ont perdu plus de 90 % de leurs effectifs. Les formations les plus épargnées sont la cariçaie évoluée (tourbière bombée évoluée) ainsi que la zone perturbée (remblais). Dans ce dernier cas, nous pouvons attribuer cette faible diminution aux bonnes capacités de déplacement et à l’abondance des espèces dans des zones proches de la plate-forme de stockage.
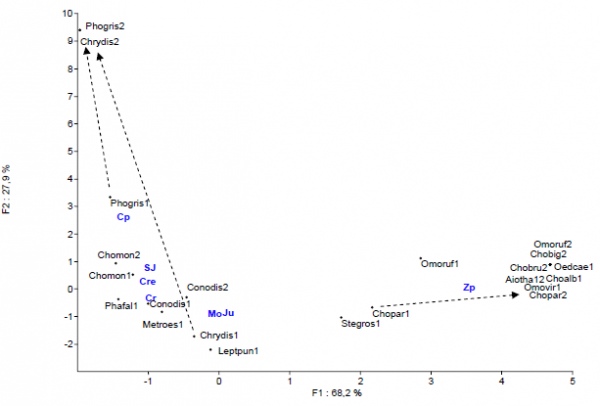
Figure 4. - Projection des relevés et espèces sur le plan F1-F2 de l’A.F.C. Code des espèces et des stations : voir les tableaux I et II. Les flèchent indiquent les changements de milieux de 3 espèces au cours de l’été.
En terme de diversité, nous notons une érosion dramatique du nombre d’espèces dans tous les milieux sauf dans la zone perturbée pour les raisons citées précédemment.
L’A.F.C. réalisée (fig. 4) permet d’opposer selon l’axe F1 les insectes de la zone perturbée (Zp) à droite de ceux des zones tourbeuses et Moliniaie-Jonçaies à gauche. Stethophyma grossum est l’espèce la plus notable présente à la fois dans les Jonçaies et dans la zone perturbée. Il faut souligner la grande homogénéité des peuplements de tourbière stricte à Carex rostrata et de zone humide de type Moliniaie-Jonçaie. Chorthippus montanus et Conocephalus discolor semblent caractériser particulièrement les Cariçaie à C. rostrata mais il faudrait étudier d’autres tourbières pour pouvoir l’affirmer. Certaines espèces présentent des variations dans leurs affinités écologiques au cours de l’été, que nous avons illustrées par des flèches sur l’A.F.C. Les plus notables concernent Chrysochraon dispar et dans une moindre mesure Pholidoptera griseoaptera qui colonisent tardivement les zones à Carex paniculata. Chorthippus parallelus quant à lui a tendance à fuir le fond tourbeux pour se cantonner à la zone perturbée à la fin de l’été, peut-être pour trouver des sites de ponte favorables.
En conclusion, nous notons une opposition entre la faune de la zone perturbée par rapport à celle que nous trouvons dans la cuvette. En effet, cette zone perturbée est en connexion, en continuité avec les milieux environnants (bords de chemins, friches, aire dénudée), ce qui facilite les échanges de populations d’espèces à large répartition et donc un maintien de la diversité. En revanche, les formations humides de la cuvette abritent une faune spécialisée dont il n’existe pas d’équivalent à proximité sachant que les formations boisées constituent des obstacles infranchissables pour la plupart des espèces.
Conclusions
Cette étude nous a permis de constater que cette zone humide est très intéressante d’un point de vue biologique car elle regroupe de nombreux milieux. Nous avons des milieux franchement tourbeux, des milieux humides plus répandus et des milieux boisés caractéristiques de la région. Parmi les milieux tourbeux, la tourbière bombée active et la tourbière boisée ont une valeur patrimoniale très importante aussi bien à l’échelle locale qu’à l’échelle européenne (MANNEVILLE, 2001).
Nos travaux n’ont toutefois pas mis en évidence d’espèces qu’il faut à tout prix préserver, mais seulement des espèces remarquables qui méritent que nous les préservions autant que faire se peut.
Cependant, il est fort possible qu’il reste des espèces intéressantes à découvrir car un travail d’inventaire, pour être le plus exhaustif possible, doit se faire sur plusieurs années.