Richesse en orthoptères et succession primaire en Haute-Vienne Richness of orthoptera and primary succession in Haute-Vienne
La richesse entomologique d’une ancienne halde de mine de la région de Bessines-sur-Gartempe est évaluée par la comparaison des connaissances acquises en Haute-Vienne sur les successions primaires. La composition de la faune et les paramètres de peuplements sont en rapport avec l’âge de la réhabilitation du site qui est de neuf ans.
The entomologic biodiversity of a former mine site in the region of Bessines-sur-Gartempe is estimated by comparison to the data acquired from primary succession studies in Haute-Vienne. Fauna composition and community parameters are in the range of what is expected for a 9 year-old site.
Introduction
Comment évaluer la richesse entomologique d’un milieu ? Peut-on qualifier de normale la diversité des insectes rencontrée dans tel endroit ? Ce sont les questions qui nous ont été posées à l’occasion de la réalisation d’inventaires dans une ancienne halde de mine de la Société AREVA, dans la commune de Bessines-sur-Gartempe, au nord de Limoges, près du hameau de Bellezane. La réponse à ces questions est moins simple qu’il n’y paraît, puisque les espèces absentes, par rapport à ce qui est attendu, sont aussi importantes que les espèces présentes.
La première idée est de comparer la composition faunistique rencontrée à ce que l’on trouve dans la même région et dans des habitats comparables. Cela nous confronte alors à l’examen d’une base de données la plus complète possible, contenant des milliers d’entrées espèces. Par exemple, il existe pour plusieurs d’ordre d’insectes du Limousin des bases de données gérées par la S.E.L. (Société Entomologique du Limousin). Une autre idée est de comptabiliser les types d’habitats, sachant qu’avec un peu d’habitude, un naturaliste peut dresser une liste d’espèces attendues par catégorie pour une région donnée. D’une manière plus formelle, c’est la méthode utilisée pour l’évaluation de la qualité des milieux, développée avec la famille des Syrphidae (Diptères) à l’échelle européenne. Les inventeurs de cette méthodologie ont conçu un logiciel qui fournit une liste prédite des espèces, que l’on compare avec la liste réelle. On pourra consulter à ce sujet le site « syrph the net : Syrphidae (Diptera) database » à l’adresse http://www.iol.ie/~millweb/ syrph/syrphid.htm. La dernière idée est la recherche d’espèces emblématiques, ou à haute valeur patrimoniale, pouvant être reconnues comme telles au niveau régional, national ou européen.
La stratégie du choix des ordres d’Insectes à prendre en compte pour réaliser des inventaires entomologiques dépend des milieux rencontrés (forêt, landes, prairies, zones humides) mais aussi de la spécialité des scientifiques. Dans la mesure où le site étudié présentait essentiellement des milieux ouverts et sans eau libre, nos efforts se sont concentrés sur la faune orthoptérologique au sens large, c’est-à-dire sur les Insectes Orthoptéroïdes (Mantoptères et Orthoptères).
L’ordre des Orthoptères comprend deux sous-ordres, à savoir les Caelifères (criquets, reconnaissables à leurs antennes courtes et la présence d’un organe de ponte court formé de 4 valves chez la femelle) et les Ensifères (Sauterelles, ou Tettigonoidea, et Grillons, ou Grylloidea, reconnaissables à leurs antennes longues et fines, et la présence d’un organe de ponte en forme de sabre plus ou moins recourbé chez la femelle, appelé oviscapte). L’intérêt des espèces appartenant aux Orthoptéroïdes est que leur identification sur le terrain est immédiate et qu’ils sont faciles à échantillonner. Ils ne se déplacent que de l’ordre de quelques mètres au cours de leur vie, soit pour une raison de dérangement, soit parce que les besoins environnementaux de l’adulte varient avec l’âge. Ces insectes forment ainsi des communautés fortement inféodées aux différentes catégories de milieux, et réagissent de manière très sensible à la hauteur et la structure de la végétation, à l’humidité du sol, et à la diversité botanique.
Des études antérieures en Limousin sur cet ordre d’insectes ont largement illustré ces notions (PICAUD et al., 2000 ; PETIT et al., 2003 et LOLIVE et PETIT, 2006). Ces communautés d’espèces sont communément utilisées pour caractériser l’état de santé d’un milieu. Nos travaux ont montré en particulier l’efficacité de leur prise en compte pour étudier les mécanismes de colonisation des terrains miniers après leur exploitation. Les résultats en sont détaillés dans la thèse de F. PICAUD (1998). Cette étude se propose donc de tester si la composition faunistique du site est en rapport avec l’âge de réhabilitation, compte tenu du calibrage déduit des travaux précédents.
Matériel et méthode
1. - Présentation du site
Le site étudié se situe à 0,8 km au NW du hameau de Bellezane sur la commune de Bessines-sur-Gartempe (Haute-Vienne) au SE du bourg (fig. 1).
Figure 1- Localisation du site
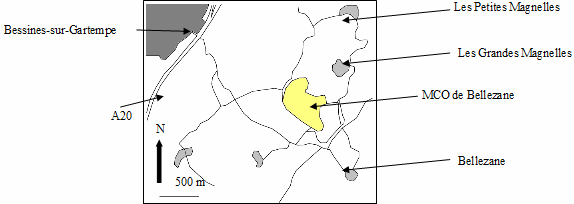
Il s’agit d’un couple de deux mines à ciel ouvert (MCO 105 et 68) réhabilitées, ayant été le siège de l’extraction de minerai d’uranium de 1983 à 1988. Avant le creusement et l’exploitation, le site était occupé par des cultures et des bois de feuillus acidiphiles à chêne pédonculé, châtaignier et bouleau, ainsi que l’atteste l’inventaire faune-flore effectué en Juin et juillet 19982 par M. A. Ghestem, Mme C. Descubes, M. D. Grafeuille avec la collaboration technique de M. C. Peyrichou (GHESTEM et al., 1982). Cet inventaire a été motivé à l’époque par le projet de creusement des deux mines à ciel ouvert.
Le site se compose de deux prairies colonisant la partie basse, entourées d’une succession de falaises et terrasses, surtout développées au niveau de la MCO 105 (fig. 2), située au SE. Le fond de cette mine à ciel ouvert est presque plan mais remonte légèrement en se dirigeant vers le nord. En revanche, le fond de la MCO 68 montre une pente également inclinée vers le sud mais plus accentuée, ce qui entraîne que la partie la plus au nord est à peine en contrebas des terrains environnants. Outre les falaises, il faut également noter des zones d’éboulis, situées à l’ouest de la partie séparant les 2 mines à ciel ouvert, à l’est de la MCO 105, la plus importante étant entourée par la boucle de l’extrême sud de la MCO 105.
2. - Relevés
Au mois de juin, nous avons noté la présence des différentes espèces d’Orthoptéroïdes arrivées au stade adulte, dans différents points du secteur. Il n’était pas cependant envisageable de travailler selon un mode quantitatif en raison de l’impossibilité de déterminer spécifiquement les larves, qui formaient alors l’énorme majorité des individus.
Les relevés standardisés d’Orthoptéroïdes ont été effectués à la fin de la saison où l’on observe le maximum de diversité, c’est-à-dire la fin du mois d’août (le 31/08/06). La hauteur de l’herbe était homogène en raison du fauchage qui a eu lieu au mois d’août. Ce fauchage a certainement entraîné une certaine mortalité des insectes, d’où une diminution des densités. Etant donnée la densité moyenne des insectes, nos relevés se sont effectués sur une surface de 9 m², délimitée par 4 objets placés sur les sommets d’un carré de 3 m de côté. Chaque relevé est choisi au hasard sur une surface homogène quant à la composition végétale, et à au moins 1 m de la formation végétale voisine, de manière à éviter les contaminations et effets de bordure. Les relevés sont séparés au moins de 10 m entre eux de manière à éviter une trop forte parenté entre individus de relevés voisins : il y a peu de chances que 2 insectes de relevés différents proviennent de la même ponte.
On parcourt la surface, en partant du bord, en attrapant tous les insectes avec un filet, puis on procède de manière concentrique jusqu’à atteindre le centre. Il est nécessaire de repasser une deuxième fois pour capturer les individus qui auraient pu échapper. Si un individu s’envole hors du carré, il est poursuivi jusqu’à la capture, de sorte que nous pouvons obtenir une liste exhaustive des espèces et une quantité fiable d’individus par espèce. Pour chacun des milieux plusieurs relevés sont effectués pour appréhender la variabilité de la composition de la faune.
L’emplacement des différents points d’échantillonnage est représenté sur la figure 2. On peut les répartir en différents milieux (BRUGEL et al., 2001) :
-
prairie mésohygrophile : D1, D2, D3, D4, K1, K2 et K3
-
prairie mésophile : B, I1, I2, I3, I4
-
prairie xérophile : F, G1, G2, G3, G4, G5, J1, J2, J3
-
prairie + rocher : J4, J5
-
chemin : partie basse (H1, H2), partie haute (H3, H4)
-
rochers sur terrasse : L1, L2, L3, L4, L5
Nous avons regroupé certains relevés pour calculer des moyennes de densités d’insectes :
B (B) ; D (D1, D2, D3, D4) ; F (F) ; G (G1, G2, G3, G4, G5) ; H1 (H1, H2) ; H3 (H3, H4) ; I (I1, I2, I3, I4) ; J (J1, J2, J3) ; J4 (J4, J5) ; K (K1, K2, K3) ; L (L1, L2, L3, L4, L5).
Figure 2 - Emplacement des relevés entomologiques. Photographie aérienne d’après le site www.geoportail.fr. D1-4 signifie que 4 relevés notés D1, D2, D3 et D4 ont été effectués.

Deux paramètres de peuplements sont ici utilisés, la diversité et la densité. L’indice de diversité retenu est l’indice de Shannon (H’). Il se calcule par la formule :
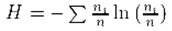
avec ni étant le nombre d’individus de l’espèce i et n le nombre d’individus au total. Les densités correspondent au nombre d’individus rapportés à 100 m². Les statistiques ont été effectuées grâce au logiciel PAST vers 1.43 (HAMMER et al., 2001).
Résultats
En tout, 32 relevés ont été effectués dans la partie prairie et sur les terrasses rocheuses. En outre, nous avons pu observer des insectes dans les ronciers et sur certaines plantes en bordure de la prairie fauchée. Par ailleurs, nous avons noté au mois de juin un certain nombre d’espèces, mais sans approche quantitative. L’ensemble des 19 espèces d’Orthoptéroïdes rencontrées est consigné dans le tableau I.
Tableau I. - Liste des espèces d’Orthoptéroïdes rencontrées dans le site.
|
Sous-ordres |
Super-familles |
Familles |
Sous-familles |
Espèces |
|
Mantoptera = Mantes |
Mantidae |
Mantinae |
Mantis religiosa |
|
|
Ensifera = Ensifères |
Grylloidea = Grillons |
Oecanthidae |
Oecanthus pellucens |
|
|
Gryllidae |
Nemobiinae |
Nemobius sylvestris |
||
|
Gryllinae |
Gryllus campestris |
|||
|
Tettigonoidea = Sauterelles |
Conocephalidae |
Conocephalinae |
Conocephalus fuscus |
|
|
Copiphorinae |
Ruspolia nitidula |
|||
|
Tettigoniidae |
Tettigoniinae |
Tettigonia viridissima |
||
|
Decticinae |
Platycleis tessellata |
|||
|
Platycleis grisea albopunctata |
||||
|
Metrioptera roeseli |
||||
|
Bradyporidae |
Ephippigerinae |
Ephippiger ephippiger |
||
|
Caelifera = Caelifères |
Acridoidea = Criquets |
Acrididae |
Calliptaminae |
Calliptamus barbarus |
|
Oedipodinae |
Aiolopus strepens |
|||
|
Oedipoda caerulescens |
||||
|
Gomphocerinae |
Euchorthippus declivus |
|||
|
Omocestus rufipes |
||||
|
Chorthippus albomarginatus |
||||
|
Chorthippus biguttulus |
||||
|
Chorthippus brunneus |
||||
|
Chorthippus parallelus |
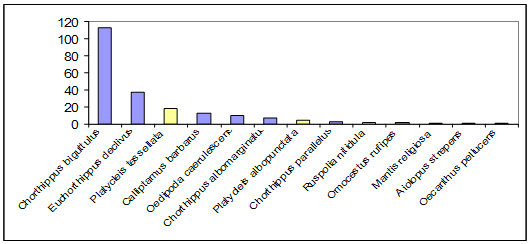
Sur l’ensemble des relevés effectués dans les prairies et terrasses rocheuses, on a 13 espèces dont l’ordre d’abondance est illustré dans la figure 3. Les deux espèces de criquets dominants sont Chorthippus biguttulus et Euchorthippus declivus, tandis que la sauterelle la plus abondante est Platycleis tessellata. Si l’on rapporte les effectifs rencontrés à des densités sur 100 m², on observe une moyenne de 90,6 individus ± 43,9. Naturellement, ces abondances relatives sont dictées par les milieux rencontrés sur le site et les affinités des espèces pour les différents milieux.
Figure 3. - Classement des espèces d’Orthoptères selon leur abondance avec en gris foncé, les Criquets et, en gris clair, les Sauterelles.
De manière à appréhender les préférences écologiques des espèces et la position relative des différents milieux, nous avons construit un tableau à double entrée, regroupant les relevés par milieux et les moyennes de densités des espèces pour effectuer une analyse statistique multivariée. En raison de la forte dominance de certaines espèces, une variante non paramétrique de l’analyse factorielle des correspondances (A.F.C.) a été appliquée, la « Detrended Correspondence Analysis », ou D.C.A. Les distances entre les projections des relevés ou des espèces ne sont pas proportionnelles aux distances réelles, comme dans une A.F.C., ce qui permet un meilleur étalement des points sur le plan factoriel F1-F2 et donc une meilleure efficacité de projection. En effet, les axes F1 et F2 représentent respectivement 51,5 % et 31,8 % de la variance du nuage de points en D.C.A. mais seulement 31,2 % et 28,4 % avec une A.F.C. L’ordination du nuage de points est illustrée à la figure 4.
Figure 4. - Ordination des espèces et des milieux sur le plan factoriel F1 F2 par D.C.A.
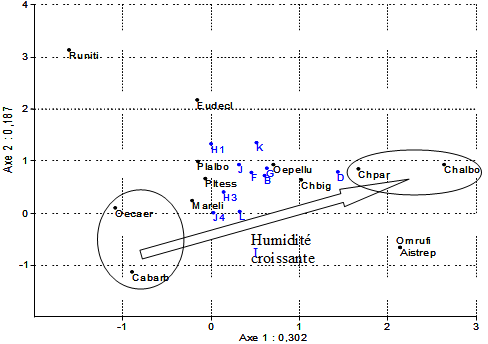
Abréviations : Pour les milieux (lettres majuscules), voir le tableau II ; Platycleis albopunctata : Plalbo ; Chorthippus biguttulus : Chbig ; Platycleis tessellata : Pltess ; Chorthippus parallelus : Chpar ; Chorthippus albomarginatus : Chalbo ; Oedipoda caerulescens : Oecaer ; Euchorthippus declivus : Eudecl ; Mantis religiosa : Mareli ; Calliptamus barbarus : Cabarb ; Ruspolia nitidula : Runiti ; Aiolopus strepens : Aistrep ; Oecanthus pellucens : Oepellu ; Omocestus rufipes : Omrufi.
Cette projection montre un gradient le long de l’axe F1, c’est-à-dire l’axe horizontal. Les espèces situées sur la gauche (Oedipoda caerulescens et Calliptamus barbarus) occupent le pôle le plus sec et leur fréquence est maximale dans 3 milieux : les terrasses rocheuses (L), la partie supérieure du chemin reliant les 2 parcelles de MCO (H3) et la prairie où affleurent des rochers (J4), dans la partie la plus élevée de la parcelle.
A l’inverse, Chorthippus parallelus et C. albomarginatus se situent à l’autre extrémité du gradient, sur la partie droite de la projection. Ces espèces caractérisent les milieux les plus humides du site, là où des joncs se mêlent aux graminées (D). Ponctuellement, on a pu rencontrer des joncs ailleurs sur le site, mais avec des taches trop restreintes pour permettre l’installation de la faune caractéristique. Les espèces les plus représentées sur le site (Chorthippus biguttulus, Euchorthippus declivus et Platycleis tessellata) occupent la partie médiane du gradient : ce ne sont pas des espèces spécialistes. Certaines espèces ont été rencontrées en trop petit nombre pour pouvoir les situer dans ce gradient, du moins par les seules données de nos échantillonnages (Omocestus rufipes, Aiolopus strepens, Ruspolia nitidula). On peut remarquer que ces 3 espèces sont les plus excentrées par rapport à l’axe du gradient.
Du point de vue des peuplements, les variations des paramètres dans les différents milieux sont synthétisées dans le tableau II.
Tableau II. - Paramètres de peuplement des Orthoptéroïdes dans les différents relevés
|
Relevés |
Densités pour 100 m² |
Shannon H’ |
|
B : prairie mésophile |
133 |
1,2 |
|
D : prairie mésohygrophile |
72,2 |
1 |
|
F : prairie xérophile |
178 |
1,2 |
|
G : prairie xérophile |
68,9 |
0,8 |
|
H1 : chemin bas |
133 |
1,6 |
|
H3 : chemin haut |
77,8 |
1,3 |
|
I : prairie mésophile |
67 |
0,998 |
|
J : prairie xérophile |
74,1 |
1,3 |
|
J4 : prairie + rochers affleurant |
55,6 |
1 |
|
K : prairie mésohygrophile |
81,5 |
0,8 |
|
L : terrasses rocheuses |
31,1 |
0,7 |
Les densités maximales caractérisent les relevés effectués en F (prairie sèche), B (prairie mésophile) et H1 (partie inférieure du chemin de séparation entre les deux parcelles), avec plus de 130 individus pour 100 m². Inversement, les densités les plus faibles (30 à 50 individus pour 100 m²) correspondent aux terrasses rocheuses (L) et à la prairie où la roche affleure (J4).
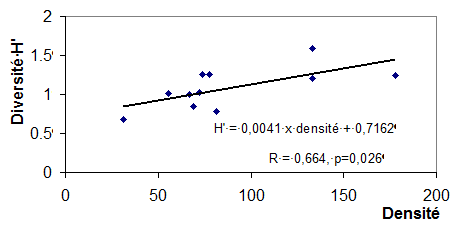
Il y a une corrélation positive significative (r = 0,664, p<0,05) entre la densité et la diversité (fig. 5), ce qui atteste de la bonne structuration des peuplements au moment de l’étude. En effet, en début de saison, on observe souvent une forte abondance de telle ou telle espèce à cause des éclosions qui sont relativement synchrones. Vers le milieu de la saison, les effectifs des populations des différentes espèces se répartissent à cause de relations interspécifiques et la diversité de chaque milieu dépend largement de la densité des espèces.
Figure 5. - Variations de la diversité de l’indice de Shannon et de la densité dans les différents milieux.
Malgré nos efforts, nous n’avons pu observer Chorthippus brunneus à la fin août alors que cette espèce était abondante à l’état adulte au mois de juin. Il est possible que les chaleurs caniculaires du mois de juillet aient accéléré son cycle ce qui a entraîné leur mort après la reproduction. Habituellement, cette espèce se rencontre jusqu’en octobre dans nos régions. Il en est de même pour la sauterelle Metrioptera roeseli dont de nombreuses larves étaient observées en juin : cette espèce précoce était absente dès la fin août. L’absence d’adultes des grillons des champs (G. campestris) est en revanche parfaitement normale dans la mesure où ces insectes se sont reproduits entre le mois de mai et le mois de juillet, puis sont morts après leur reproduction. Ce sont les larves qui passeront la mauvaise saison, conformément au cycle de l’espèce.
Il est intéressant de rappeler que les ronciers abritent 2 espèces de sauterelles, aisément repérables grâce à leur chant, à savoir Ephippiger ephippiger et Tettigonia viridissima. Cette dernière avait été notée en grande abondance (environ un individu pour 1 ou 2 m²) dans les hautes herbes de la partie sud du site, à la fin du mois de juin. En raison de la fauche et des travaux conduits au cours de l’été, cette espèce s’est réfugiée dans les ronciers à la fin du mois d’août. E. ephippiger, quant à elle, est une espèce plus tardive et aucun adulte n’était présent à la fin juin.
Discussion
La liste des 20 espèces d’Orthoptéroïdes rencontrées est conséquente par rapport à la soixantaine d’espèces que compte la région Limousin. Cette diversité augure d’une qualité du milieu satisfaisante dans l’ensemble du site. Cependant il convient d’avoir un avis critique sur la signification de la présence ou de l’absence de certaines espèces. Les règles écologiques qui conditionnent la composition faunistique sont simples et découlent de la conjugaison de la diversité des milieux (comprenant la surface des différents microhabitats) et l’âge de la restauration (notion non indépendante de la facilité d’accès par rapport aux alentours du site). Dans la thèse de F. PICAUD (1998), synthétisée en partie dans son article (PICAUD et PETIT, 2006), l’analyse de sites miniers de Haute-Vienne dont la restauration couvrait un éventail compris entre 0 à 9 ans, a permis d’établir un profil d’espèces associées aux stades jeunes (avant 2,5 années), moyens et tardifs (plus de 4,5 années). Les résultats mettent en avant un aspect qualitatif : les espèces qui arrivent en premier sur un site ne sont pas statistiquement les mêmes que celles qui colonisent en dernier. Les 3 groupes d’espèces, appelés des sères, sont définis dans le tableau III :
Tableau III. - Composition des 3 sères d’Orthoptères colonisant les sites miniers restaurés de Haute-Vienne. Les espèces rencontrées sur le site de Bellezane sont indiquées en gras.
Sère 1 : Aiolopus thalassinus (en mélange avec A. strepens), Chorthippus brunneus, Oedipoda caerulescens et Calliptamus italicus (en mélange avec C. barbarus) ;
Sère 2 : Chorthippus biguttulus, Omocestus rufipes, Stethophyma grossum, Stenobothrus stigmaticus, Nemobius sylvestris, Gryllus campestris, Ephippiger ephippiger et Platycleis albopunctata grisea ;
Sère 3 : Chorthippus dorsatus, Chorthippus montanus, Chorthippus parallelus, Euchorthippus declivus, Conocephalus discolor, Platycleis tessellata, Phaneroptera falcata et Tettigonia viridissima.
La place de Chorthippus albomarginatus est incertaine et se situe entre les sères 2 et 3.
Le tableau III montre clairement que les espèces du site de Bellezane se répartissent dans les 3 sères. Cela signifie donc que la composition de ce site correspond à un âge de réhabilitation de plus de 4,5 années. Les espèces pionnières (sère 1), à fort pouvoir de dispersion, n’ont pas disparu mais se sont réfugiées au niveau des prairies les plus xérophiles, les prairies où le rocher affleure et les terrasses rocheuses. Ces milieux sont ceux qui ressemblent le plus aux terrains jeunes, caractérisés par une végétation dispersée et la présence de sol nu. Les espèces intermédiaires (sère 2) se maintiennent à la faveur du fauchage annuel de l’herbe, comme pour C. biguttulus, O. rufipes ou G. campestris. Les espèces tardives (sère 3), quant à elles, ont généralement un faible pouvoir de dispersion (PICAUD et PETIT, 2007) mais une fécondité particulière caractérisée par des oeufs de forte taille.
Ces résultats observés sont parfaitement cohérents avec l’âge réel de la réhabilitation, c'est-à-dire 9 ans. Un autre résultat aurait évidemment signifié soit un problème d’accessibilité à la mine, soit une source de perturbation, comme une pollution ou des travaux mécaniques.
Du point de vue quantitatif, les paramètres de peuplement sont également instructifs. On peut comparer les densités et diversités observées sur le site aux chiffres mesurés par PICAUD et PETIT (2006), pour les mois d’août et de septembre. Les valeurs observées sur la partie prairie (tous les relevés sauf les L, sur terrasse rocheuse) sont rassemblées dans le tableau IV, de manière à faciliter les analyses.
Tableau IV. - Paramètres de peuplement sur le site et sur le calibrage de PICAUD et PETIT (2006)
|
Mois |
Sites miniers, prairies > 4,5 ans |
Prairies de fauche des alentours des mines |
Site MCO 68+105 (Fin août) |
|
|
Densité /100 m² |
Août |
285 |
98,1 |
98,4 |
|
Septembre |
64 |
41,7 |
||
|
Diversité H’ |
Août |
2 |
2,2 |
1,13 |
|
Septembre |
1,9 |
1,6 |
La densité moyenne au niveau de la prairie de fauche dans le site étudié est proche de 100, ce qui est intermédiaire entre les valeurs d’août et de septembre dans les sites réhabilités depuis plus de 4,5 ans. En revanche, la diversité mesurée par l’indice de Shannon est de l’ordre de 1,13, ce qui est plus faible que les valeurs trouvées en août et septembre pour des sites réhabilités depuis plus de 4,5 ans. Si on compare maintenant ces paramètres avec ceux des alentours des sites miniers, dans les prairies, on a une fois de plus des chiffres comparables pour ce qui est de la densité, mais inférieurs pour ce qui est de la diversité.
Globalement, pour ce qui est de la diversité, on a une contradiction entre les chiffres trouvés par rapport au nombre global d’espèces sur le site et la moyenne des valeurs trouvées par relevé. Ce nombre global est très satisfaisant alors que par relevé, la diversité est insuffisante par rapport à ce que l’on pouvait attendre.