Les capacités migratoires de Galba truncatula et d’Omphiscola glabra (Lymnaeidae) dans des ruisseaux sur sol acides et leurs conséquences sur la transmission de la fasciolose The ability of Galba truncatula and Omphiscola glabra (Lymnaeidae) to upstream migrate in brooks on acid soils
Daniel RONDELAUD ,
Philippe VIGNOLES ,
Gilles DREYFUSS
Philippe Hourdin
Comme les gîtes permanents de Galba truncatula et d’Omphiscola glabra sont situés à une distance de 3 à 67 m des cressonnières naturelles dans la région du Limousin (situées généralement au point d’émergence de sources permanentes), des investigations ont été réalisées pendant deux années sur 12 rigoles de drainage, alimentées chacune par une source permanente, afin d’étudier la migration hivernale de ces limnées en amont vers les sources et de déterminer les conséquences de ces migrations sur la contamination des cressonnières par le parasite. Les migrations des mollusques ont été suivies sur des distances de 30 ou 60 m entre les points où ils ont été introduits dans les rigoles et les cressonnières situées à l’émergence des sources. Sur les effectifs de départ, seuls 15,8 % des G. truncatula et 12,8 % des O. glabra ont colonisé les sources. Les O. glabra migrent plus vite que G. truncatula car les premières limnées ont été trouvées dans les sources à la 7e semaine alors que les secondes n’ont été recensées qu’à la 10e semaine. Malgré cette migration plus rapide d’O. glabra, les infestations naturelles avec F. hepatica sont plus nombreuses chez G. truncatula, avec des prévalences significativement plus élevées chez les F1 de G. truncatula que chez ceux de l’autre limnée. Le faible nombre de mollusques capables de migrer jusqu’aux cressonnières en amont peut s’expliquer par l’existence d’un état physiologique particulier, probablement lié à un retard dans le développement de l’activité reproductrice. Les infestations naturelles avec F. hepatica, relevées dans ces cressonnières, soulignent l’importance de la présence des mollusques hôtes pour qu’il y ait contamination du cresson par le parasite.
As the reservoir habitats of Galba truncatula and those of Omphiscola glabra in the region of Limousin (central France) were located at a distance of 3 to 67 m from natural watercress beds (usually set around the sources of permanent springs), field investigations in 12 open drainage furrows, each supplied by a permanent spring, were carried out over two years to study the upstream migration of snails in winter from their habitats towards the beds and to determine the consequences of these snail migrations on the contamination of watercress beds with Fasciola hepatica. Snail migrations were bi-weekly followed on 30-m or 60-m distances, taking place between the points of snail introduction in furrows and the upstream beds. Out of the numbers of snails introduced at the onset of the experiment, 15.8 % of G. truncatula and 12.8 % of O. glabra only colonized spring sources. O. glabra migrated quicker than G. truncatula, as the presence of the former snails was noted at week 7 in sources, while the latter were only found at week 10. In spite of the quicker upstream migrations of O. glabra, the natural infections of snails with F. hepatica were greater in G. truncatula and the corresponding prevalences were significantly higher in the F1 of this lymnaeid species than in those of O. glabra. The low number of snails able to upstream migrate up to watercress beds might be explained by the existence of a particular physiological state for these snails, probably linked with a delay in the development of their reproductive activity. The natural infections of snails with F. hepatica, found in these beds, underlined the importance of the presence of snail hosts for watercress contamination by the parasite.
Introduction
On sait depuis longtemps que les Gastéropodes d’eau douce sont capables de remonter les rivières et les ruisseaux à contre-courant (rhéotropisme) pour aller vers les sources. Les deux exemples les plus connus sont ceux de Potamopyrgus antipodarum (Prosobranche) qui a colonisé tout le réseau hydrographique français à partir des estuaires (Real, 1973 par exemple) et de Physella acuta (Pulmoné) qui peut remonter rapidement un ruisseau ou un système de drainage superficiel lorsque les conditions s’y prêtent (Vareille et al., 1996). Les capacités des limnées pour effectuer un tel rhéotropisme ont été moins étudiées. Galba truncatula est capable de coloniser de nouvelles aires à partir de ses habitats permanents (Taylor, 1965) et cette aptitude serait essentiellement liée aux déplacements du mollusque à contre-courant (Hohorst, 1969 ; Moens, 1982). Dans les prairies de la Haute-Vienne, l’espèce peut envahir un réseau de drainage superficiel (dépourvu de sources permanentes) en trois années. Les distances parcourues par rhéotropisme sont plus importantes de mars à juin (lorsque les conditions sont favorables) alors qu’elles sont faibles de novembre à janvier et généralement nulles en août et septembre (Rondelaud, 1983). Dans le cas de l’autre limnée : Omphiscola glabra, la revue de la littérature ne montre pas d’étude sur ce point particulier.
Comme ces deux limnées vivent l’une à côté de l’autre sur la même rigole de drainage dans les sols acides de la Haute-Vienne (Vareille-Morel et al., 1999), elles participent donc toutes les deux à la remontée hivernale et printanière vers les sources. Lorsque ces dernières sont permanentes, elles sont souvent peuplées par du cresson de fontaine (Nasturtium officinale) si bien qu’il y a des risques de distomatose à Fasciola hepatica. En effet, les deux limnées précitées sont connues pour intervenir, à des degrés divers, comme hôtes intermédiaires dans le cycle de ce parasite et la présence d’une espèce, de l’autre, des deux ou, au contraire, leur absence dans une cressonnière donnée retentit sur la contamination de celle-ci par le parasite (Rondelaud et al., 2005). Devant ce problème particulier, il était utile de connaître les capacités de ces deux limnées pour migrer en amont dans des rigoles de drainage alimentées en permanence par des sources. C’est la raison pour laquelle nous nous somme posé les trois questions suivantes : quelle est la distance que chaque espèce de limnée parcourt lors de son déplacement à contre-courant vers les sources ? combien de mollusques peuvent atteindre le point d’eau dans lequel pousse le cresson ? et quelles sont les conséquences à terme d’une telle migration sur la contamination d’une cressonnière par F. hepatica ? Pour répondre à cette problématique, une étude expérimentale a été réalisée en 2003 et 2004 dans le milieu naturel en plaçant des groupes de G. truncatula ou d’O. glabra à des distances connues de sources permanentes situées en amont et en étudiant les migrations de ces mollusques pendant les six premiers mois de chaque année. Des investigations parasitologiques ont également été effectuées en disséquant les survivants et leur descendance à la recherche des formes larvaires de F. hepatica.
Matériel et méthodes
Douze rigoles de drainage superficiel, situées dans le nord de la Haute-Vienne et ali-mentées chacune par une source permanente, sont concernées par cette étude. Elles ont été choisies en raison de l’absence des limnées depuis de nombreuses années alors qu’une cressonnière est présente dans la source. Leur longueur varie de 75 à 127 m et leur largeur est de 45 cm en moyenne. Leur pente est de 1,2-1,4 %. La vitesse de l’eau fluctue de 16 à 24 cm/sec selon les rigoles en janvier et diminue ensuite pour n’être plus que de 3 à 4 cm/sec en juin.
Tableau I. Les différentes étapes de l’expérience réalisée pour suivre les migrations en amont de Galba truncatula et d’Omphiscola glabra en 2003 et 2004. *, G. truncatula : 4-5 mm de hauteur ; O. glabra, 9-11 mm.
|
Année |
Témoins ou Expérimentés |
Nombre de rigoles (effectif des limnées * par rigole) |
Distance à parcourir jusqu’à la cressonnière |
|
2003 |
Expérimentés |
8 (40 Galba truncatula + 40 Omphiscola glabra) |
30 m |
|
2004 |
Témoins |
2 pour G. truncatula (80 limnées), |
60 m |
|
2 pour O. glabra (80 limnées) |
|||
|
Expérimentés |
8 (40 G. truncatula + 40 O. glabra) |
Le tableau I précise les différentes étapes de l’expérience. Cette dernière a été réalisée pendant deux années, en 2003 et 2004. Au début de janvier 2003, un groupe mixte comprenant 40 G. truncatula transhivernants, hauts de 4 à 6 mm et 40 O. glabra mesurant 9 à 11 mm a été introduit dans chacune des huit rigoles (voir le tableau I), à une distance de 30 m de la cressonnière située en amont. Les hauteurs de G. truncatula et celles de O. glabra ont été choisies en fonction des résultats d’une étude préliminaire (Rondelaud et al., 2005) car ce sont les dimensions que la plupart des mollusques retrouvés dans les cressonnières entre 1999 et 2002 présentent au printemps. L’expérience a été renouvelée au début de janvier 2004 en utilisant les mêmes rigoles et le même protocole mais la distance que les mollusques doivent parcourir est de 60 m. De plus, quatre rigoles témoins ont été suivies en introduisant un groupe de 80 G. truncatula dans chacune des deux premières stations et un groupe de 80 O. glabra dans les deux autres.
Des investigations bihebdomadaires ont été réalisées par la suite pour suivre la migration de ces mollusques en amont vers la cressonnière, pour déterminer le nombre de mollusques qui colonisent chaque source et pour mesurer le temps (en semaines) que mettent les limnées les plus rapides pour parcourir 30 ou 60 m. La distance moyenne parcourue chaque semaine par l’ensemble du groupe a été également calculée. Chaque année, à la fin du mois de juin, les mollusques transhivernants (génération parentale) et leurs descendants (génération F1) ont été récoltés dans chaque cressonnière ou dans les alentours immédiats avant d’être transportés au laboratoire dans des conditions isothermes et d’être disséqués sous un stéréomicroscope pour y rechercher d’éventuelles formes larvaires de F. hepatica.
Le premier paramètre étudié est le nombre des mollusques présents dans chaque tranche de rigole (de 15 m chacune) et dans la source qui alimente la cressonnière. Cet effectif est déterminé pour chaque espèce de limnée (G. truncatula, ou O. glabra) et pour chaque semaine durant la migration en amont des mollusques (soit 13 semaines au maximum). Les quatre paramètres suivants ne concernent que les limnées les plus rapides et se rapportent i) à leur nombre pour chaque année et chaque type de rigole (à peuplement mixte ou à population monospécifique), ii) à la hauteur de coquille que ces limnées présentent en arrivant dans la source, iii) à la durée de la migration (en semaines) depuis le point d’introduction des limnées au départ de l’expérience) jusqu’à la source, et iv) à la distance parcourue tous les 3,5 jours par les mollusques (3 par rigole). Les valeurs individuelles, recueillies pour chacun de ces paramètres, sont ramenées à une moyenne, encadrée d’un écart type, en tenant compte de l’espèce de la limnée, de l’année et du type de rigole (pour la seule année 2004). Enfin, la contamination des cressonnières par F. hepatica a été suivie au mois de juin de chaque année en récoltant les mollusques présents dans ces sites (parents et mollusques F1) et en les disséquant sous loupe binoculaire à la recherche d’éventuelles formes larvaires. Les moyennes obtenues pour les cinq derniers paramètres ont été comparées par une analyse de variance à un seul facteur (Stat-Itcf, 1988) en tenant compte de l’année d’étude et de l’espèce de la limnée.
Résultats
1. Les migrations des mollusques.
Tous les mollusques ne participent pas à la migration printanière. A titre d’exemple, nous avons présenté, sur le tableau II (page suivante), la répartition des 320 mollusques de chaque espèce dans les huit rigoles qui ont reçu des groupes mixtes de mollusques en 2004. Si l’on considère les chiffres à la 13e semaine, on constate que 54,3 % des G. truncatula et 76,6 % des O. glabra sont restées dans les tranches de rigole qui touchent le point d’introduction des limnées au départ de l’expérience (de - 15 m à + 30 m). Les autres ont participé à la migration mais seules quelques limnées, à savoir 15,8 % des G. truncatula et 12,8 % des O. glabra ont effectivement colonisé les sources. Les O. glabra se déplacent, d’autre part, plus vite que G. truncatula car les premières limnées ont été trouvées dans les sources à la 7e semaine alors que les secondes n’ont été recensées qu’à la 10e semaine.
Au vu de ces résultats, les paramètres n’ont été déterminés que pour les limnées les plus rapides (les trois premières qui ont colonisé chaque cressonnière). Le tableau III présente les valeurs pour trois de ces variables. Le nombre total d’O. glabra par cressonnière est significativement plus élevé (F = 6.83, P < 5 %) en 2003 que celui noté pour G. truncatula alors qu’en 2004, il n’y a pas de différence significative entre les deux espèces.
Tableau II. La répartition des deux limnées au cours des 13 semaines de l’expérience dans les huit rigoles colonisées chacune par un groupe de 80 mollusques (40 Galba truncatula + 40 Omphiscola glabra). *, le point 0 dans chaque rigole correspond au point d’introduction des mollusques au départ de l’expérience. †, chiffres cumulatifs. Les pourcentages à la 13e semaine ont été calculés par rapport à l’effectif des mollusques retrouvés en vie.
|
Semaine (2004) |
Longueur de la rigole * |
Non retrouvés ou morts † |
|||||
|
-15 m/0 |
0/15 m |
15,1/30 m |
30,1/45 m |
45,1/60 m |
Source |
||
|
Nombre de Galba truncatula transhivernants |
|||||||
|
1 |
0 |
320 |
0 |
0 |
0 |
0 |
0 |
|
4 |
31 |
228 |
54 |
0 |
0 |
0 |
7 |
|
7 |
43 |
170 |
72 |
15 |
1 |
0 |
19 |
|
10 |
27 |
95 |
104 |
27 |
14 |
10 |
43 |
|
13 |
6 (3,7 %) |
41 (25,0 %) |
42 (25,6 %) |
31 (18,9 %) |
18 (11,0 %) |
26 (15,8 %) |
156 |
|
Nombre d’Omphiscola glabra transhivernants |
|||||||
|
1 |
0 |
320 |
0 |
0 |
0 |
0 |
0 |
|
4 |
11 |
178 |
73 |
26 |
0 |
0 |
32 |
|
7 |
5 |
126 |
49 |
32 |
14 |
20 |
74 |
|
10 |
1 |
61 |
36 |
15 |
9 |
29 |
140 |
|
13 |
1 (1,2 %) |
43 (50,0 %) |
21 (24,4 %) |
7 (8,1 %) |
3 (3,5 %) |
11 (12,8 %) |
234 |
Tableau III. Valeurs moyennes et écarts types pour trois paramètres se rapportant aux limnées qui ont colonisé en premier les sources. (T), rigoles témoins. Hauteur des mollusques au départ : 4,9 ± 0,5 mm (Galba truncatula), 10,1 ± 1,7 mm (Omphiscola glabra).
|
Paramètres |
Longueur de rigole et année |
Galba truncatula |
Omphiscola glabra |
|
Nombre de limnées atteignant les sources en premier |
30 m, 2003 60 m, 2004 60 m, 2004 (T) |
3,8 ± 1,4 1,2 ± 0,9 1,5 ± 0,8 |
6,8 ± 2,1 2,4 ± 1,3 3,0 ± 0,9 |
|
Durée de la migration (en semaines) |
30 m, 2003 60 m, 2004 60 m, 2004 (T) |
5,3 ± 1,1 9,6 ± 2,6 9,1 ± 1,8 |
3,7 ± 0,8 5,9 ± 1,1 6,3 ± 1,3 |
|
Hauteur des limnées atteignant les sources en premier (en mm) |
30 m, 2003 60 m, 2004 60 m, 2004 (T) |
6,5 ± 1,7 8,2 ± 2,0 8,1 ± 1,7 |
13,2 ± 2,1 15,6 ± 2,3 16,0 ± 2,1 |
Fig. 1a-d. Moyennes et écarts types pour les distances parcourues par les limnées les plus rapides dans les rigoles à peuplement mixte (2003 : 1a, et 2004 : 1b) et dans celles avec une seule espèce de mollusque (1c et 1d). Deux relevés sont effectués chaque semaine pendant les dix semaines de la migration en amont. Galba truncatula (histogrammes en blanc) et Omphiscola glabra (en hachurés).
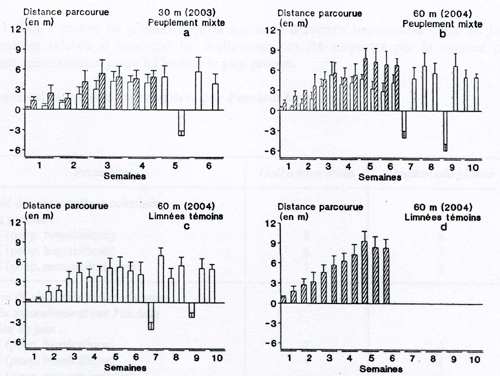
Les durées de migration pour parcourir 30 et 60 m sont significativement plus faibles (2003 : F =. 5,63, P < 5 % ; 2004 : F = 7,82, P < 5 %) pour O. glabra que pour l’autre limnée. Quant à la hauteur des mollusques, elle ne présente pas de variation nette entre les moyennes recueillies en 2003 et celles notées en 2004, quelle que soit l’espèce de la limnée. La comparaison des résultats obtenus pour les rigoles témoins (à peuplement monospécifique) avec les données fournies par les groupes mixtes ne montre pas de différences significatives pour chaque espèce de limnée prise isolément, quel que soit le paramètre considéré. Même si l’effectif des limnées pour une espèce donnée est plus important (80 mollusques au lieu de 40) au départ de l’expérience, le nombre de celles qui atteignent la cressonnière est identique, ce qui indique que l’effectif des limnées qui migrent en amont est indépendant de l’abondance de la population.
La figure 1 montre les distances que les limnées les plus rapides ont parcourues en 2003 et 2004, aussi bien dans les rigoles à peuplement mixte que dans celles avec une seule espèce de mollusque (témoins). Les valeurs augmentent au cours des trois premières semaines et passent par un palier chez G. truncatula jusqu’à la fin de la migration (à la 10e semaine). Par contre, chez O. glabra, l’accroissement des distances s’observe jusqu’à la 5e semaine (Fig. 1b, d) ou au contraire laisser la place à un palier à partir de la 3e semaine (Fig. 1a). A l’exception de 2003 pour laquelle aucune différence significative n’a été retrouvée entre les longueurs parcourues par les deux limnées, les distances recensées lors de la migration en amont des deux espèces à partir de la 4e semaines sont significativement plus importantes (F = 15,1 ; P < 1 %) chez O. glabra que chez G. truncatula.
Les valeurs négatives notées pour G. truncatula sur les trois premiers graphes de la figure 1 sont à mettre en relation avec la survenue d’averses importantes dans les jours précédant les relevés si bien que les mollusques ont été emportés par le courant plus important, généralement jusqu’à l’herbier le plus proche.
2. La contamination des cressonnières par Fasciola hepatica.
Tableau IV. La contamination des cressonnières naturelles (situées à l’émergence des sources) par Fasciola hepatica.
|
Paramètres |
Galba truncatula |
Omphiscola glabra |
|
Nombre de cressonnières colonisées par les parents : - 2003 (peup. bispécifique) - 2004 (peup. bispécifique) - 2004 (peup. monospécifique) |
8 6 1 |
8 7 2 |
|
Nombre de sites avec des mollusques infestés naturellement par Fasciola hepatica en juin : - 2003 (peup. bispécifique) - 2004 (peup. bispécifique) - 2004 (peup. monospécifique) |
7 4 1 |
3 1 0 |
|
Prévalence moyenne de l’infestation par F. hepatica (et écart type) dans la génération F1 en juin : - 2003 (peup. bispécifique) - 2004 (peup. bispécifique) - 2004 (peup. monospécifique) |
4,3 % (0,9) 3,5 % (0,7) 2,9 % |
1,3 % (1,0) 1,1 % 0 % |
Malgré la migration plus rapide d’O. glabra vers les cressonnières situées en amont, les investigations réalisées dans les cressonnières au mois de juin 2003 (ou 2004) ont montré que les infestations naturelles étaient plus nombreuses chez G. truncatula que chez O. glabra, aussi bien dans les rigoles à peuplement mixte (7 populations en 2003 au lieu de 3, 4 populations en 2004 au lieu d’une seule) que dans les stations témoins (1 population de G. truncatula et aucune pour O. glabra). La dissection des survivants de la génération parentale (celle qui a migré en amont) n’a pas permis de trouver de formes larvaires de F. hepatica, quelles que soient l’année et l’espèce de la limnée. Par contre, des individus parasités ont été retrouvés dans leurs descendants (génération F1) et les prévalences obtenues sont significativement plus importantes (F = 20,17 ; P < 1 %) chez G. truncatula que chez l’autre limnée, quelle que soit l’année d’étude pour les rigoles à peuplement mixte.
Discussion.
Nos résultats sur les migrations des limnées vers l’amont d’un réseau hydrographique sont en accord avec les observations que l’un d’entre nous (Rondelaud 1983) a obtenues dans plusieurs prairies marécageuses de la Haute-Vienne lors de la recolonisation du réseau de drainage superficiel par des G. truncatula provenant d’habitats situés plus en aval. En particulier, les distances parcourues s’inscrivent dans le même ordre de grandeur, qu’il y ait une source permanente ou non à l’extrémité de la rigole de drainage. Ce dernier résultat est intéressant car il démontre que la vitesse du courant n’interviendrait que faiblement sur l’amplitude des distances hebdomadaires parcourues par les limnées. L’accroissement de ces dernières au fur et à mesure des semaines serait donc à rapporter à la seule température de la nappe d’eau (celle-ci s’accroît de janvier à mars dans les cours d’eau de la région), comme Boray (1969) l’a déjà rapporté en Australie lors de la colonisation de canaux par une autre limnée, Austropeplea tomentosa.
Deux données appellent, cependant, des commentaires particuliers :
- 1) La première se rapporte aux nombres des limnées qui sont capables de peupler la cressonnière située en amont, au terme d’une migration de plusieurs semaines. Ces effectifs restent faibles : 15,8 % des G. truncatula et 12,8 % des O. glabra. Dans ces conditions, on peut se demander pourquoi ces faibles nombres de limnées migrent vers l’amont alors que les autres membres du groupe restent dans le voisinage du lieu d’introduction ou ne présentent que des migrations plus faibles. La densité des mollusques introduits, à savoir 80 mollusques par rigole en un seul endroit, et la recherche de la nourriture ne peuvent, à notre avis, être les facteurs déterminants qui induisent une migration d’une telle amplitude chez quelques mollusques. L’hypothèse la plus logique serait d’admettre l’existence d’un état physiologique particulier chez les limnées migratrices, probablement lié à leur activité reproductrice. En effet, les pontes des G. truncatula transhivernants s’observent à partir de la 5e semaine d’expérience aux alentours des lieux d’introduction dans les rigoles alors qu’elles ne sont déposées qu’au cours de la 9e semaine aux environs des cressonnières. Si l’on retient cette hypothèse comme valide, il faut alors admettre que l’énergie dépensée par les mollusques pour remonter à contre-courant le cours d’eau se traduirait par un retard dans le développement de leur activité reproductrice.
- 2) Les déplacements d’O. glabra sont plus rapides que ceux de l’autre limnée et ce résultat est confirmé par l’amplitude des distances hebdomadaires parcourues. Pour expliquer ce fait, le premier argument est de rapporter cette différence à la hauteur de coquille des deux espèces car les individus utilisés pour cette expérience mesuraient 4 à 6 mm chez G. truncatula et 9 à 11 mm chez O. glabra. Cependant, cette interprétation ne peut tout expliquer, en particulier l’accroissement net des valeurs obtenues pour cette dernière limnée au cours des 5e et 6e semaines. Dans ces conditions, il faut admettre l’intervention d’un autre facteur et ce dernier pourrait être une nage à la surface de l’eau car ce mode de déplacement est fréquent chez O. glabra au cours de la période précitée. Dans ces conditions, cette nage en surface permettrait à l’espèce d’aller plus vite dans sa remontée en amont, surtout si le vent local souffle dans le même sens que les déplacements des mollusques.
Les infestations naturelles avec F. hepatica, constatées chez les F1 des deux limnées, ont été relevées dans des cressonnières pour lesquelles aucun cas de contamination avec le parasite n’avait été relevé au cours des trente dernières années. Il est donc important que les mollusques hôtes soient présents dans le site pour qu’il y ait contamination, car les hôtes définitifs du parasite, en particulier des Lagomorphes, sont présents de manière régulière autour des cressonnières et présentent des prévalences élevées lors d’infestations naturelles avec le parasite (42 %, par exemple, pour Oryctolagus cuniculus d’après Rondelaud et al., 2001). Les pourcentages plus élevés dans le cas de G. truncatula, peuvent s’expliquer facilement par le fait que les prévalences des infestations naturelles sont toujours plus faibles chez O. glabra lorsque les deux espèces de limnées vivent dans la même rigole (Abrous et al., 1999, 2000). En revanche, l’absence de formes larvaires chez les mollusques survivants de la génération parentale ne peut s’interpréter qu’à partir des données de Gold et Goldberg (1979). D’après ces auteurs, la sensibilité de l’espèce à l’infestation avec F. hepatica diminue lorsque la taille du mollusque augmente.